
20 世纪 40 年代第一个抗生素药品—青霉素的出现开创了人类治疗疾病的新时代,经过 70 余年的不断完善,各种抗生素相继问世增强了人类对传染性疾病的抵抗力。然而随着抗生素药物的广泛和大量使用,细菌的耐药问题也日趋严重。细菌抗生素耐药能力的不断上升以及耐药相关基因在不同病原菌间的传播已经成为临床感染性疾病治疗的难题。对当前几乎所有临床用抗生素均产生耐药性的“超级细菌”越来越频繁的出现更引发了社会对抗生素滥用和细菌耐药问题的担扰,这一问题已引起多国政府和卫生部门的高度重视。实际上近 30 年来,细菌耐药性的研究一直是医学微生物研究的热点,而细菌的耐药性研究先后导致了耐药质粒、整合子、整合性接合元件等可移动遗传元件( mobile genetic elements,MGE)的发现,它们是细菌耐药基因以及其它细菌适应性功能基因水平转移的主要载体。
1、 SXT /R391 元件的发现
20 世纪 90 年代以前,世界范围内流行的霍乱均由01 群霍乱弧菌( Vibrio cholorae) 引起,1992 年从印度和孟加拉国爆发的霍乱迅速波及到周边十几个国家和地区,引发此次霍乱爆发的 O139 群霍乱弧菌除了代表一种新的血清群外,另一个显著特点是对磺胺甲基异恶唑、甲氧苄啶、链霉素、氯霉素耐药。O139 群霍乱弧菌的典型株 MO10 的耐药性是由其染色体上一种可自我转移的遗传元件 SXT 介导的,在此之前并未在霍乱弧菌中发现此类元件。后来发现 1972 年分离自南非的雷氏普罗威登斯菌( Providenciarettgeri) 所携带的“F 因子”实质上是一种与 STX 密切相关的可自我转移的遗传元件,命名为 R391,R391 赋予宿主卡那霉素耐药性及重金属离子 Hg2 +抗性。SXT、R391 以及后来发现的多个相关的元件具有高度类似的保守骨架结构,以及固定的染色体插入/重组位点,因此被划分为同一个SXT / R391 家族。2011 年,Mata 等发现: 19 个对大环内酯类抗生素耐药的奇异变形杆菌 ( Proteusmirabilis) 临床株中,有 7 株( 37% ) 携带 SXT / R391,这是第一次报道 SXT/R391 广泛存在于奇异变形杆菌临床株中,并且耐药基因 blaCMY-2位于 SXT/R391 中。
截止到2012 年,在临床和环境的细菌株中超过 40 个SXT / R391 被发现。SXT/R391 是整合性接合元件( Integrating conjugative elements,ICE) 家族中一个类别,ICE 是一类可自我转移的遗传元件,它们可自主从宿主染色体剪切并通过宿主与受体菌接合形成的通道进入受体菌,然后整合到受体菌的染色体上。SXT/R391 家族目前已经 ICE 元件中多样性最为丰富、成员最多的一个家族。
2、 SXT /R391 结构、基因功能
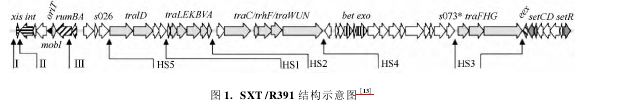
通过对 13 个不同来源的 SXT/R391 。完全序列的比较以及其它 ICE 的认识,Wozniak 等确定了SXT / R391 保守的核心骨架结构、部分保守基因功能以及外源 DNA 的插入位点( 图 1) 。SXT/R391 保守的核心区域主要涉及 SXT/R391 的整合/剪切、接合转移、元件表达调节。整合酶 Int 负责 SXT/R391 位点特异性整合; xis 的编码产物为重组定向因子 Xis。在 Xis 协助下,Int 也负责 SXT/R391 位点特异性剪切; RumAB 负责趋错 DNA 的修复。
oriT 为 SXT / R391 转移的起始位点,为保守的顺式作用元件,长 299bp,位于保守的基因 s003 与 mobI之间,在解旋酶 TraI 的作用下,环形的双链 DNA 在此处形成缺口,并最终形成可以通过接合孔隙的单链 DNA。位于 oriT 下游的 mobI,为转移过程必需基因,其功能与 oriT 识别有关。一系列 Tra基因则涉及 SXT/R391 接合和转移的过程,包括DNA 加工以及 DNA 输出的交配孔道形成、单链DNA 运输等。Bet、Exo 负责 SXT/R391 间的重组,traG、eex 则为编码排斥系统的基因,用于阻止相同拷贝的 SXT/R391 在同一个宿主细胞中出现。SXT/R391 核心区还包括一些未知功能基因,研究发现即使所有这些未知功能基因全部缺失也不会影响 SXT/R391 的转移频率,因此推测核心区这部分未知功能的基因可能与增强 SXT/R391 的适应性有关SXT / R391 有 5 个区域为可变的外源 DNA 经常插入的区域,称为热点区域 HS1-HS5( 图 1) 。
此外,在一些 SXT/R391 也发现了其它的插入外源DNA 的区域,目前已经鉴定出 4 个这样的区域,命名为 VRⅠ-VR Ⅳ( 图 1)。这些可变区在每一个元件中的总长度可以达到 30 -60 Kb。目前发现 SXT/R391 可变区基因的常见的编码产物主要负责宿 主对抗生素的耐药性、重金属离子抗性、生物膜形成和细菌运动能力的调节,另外也编码毒素-抗毒素系统以阻止 ICE 从宿主丢失。此外,一些 SXT/R391 的可变区还具有其它一些功能,例如: 编码限制性修饰系统、解旋酶、核酸内切酶,它们的作用可能是阻碍外源 DNA 的入侵或者保持 ICE 转移期间的完整性。Balado 等通过噬菌体感染实验证实了 2 株不同种弧菌中的SXT / R391ICEVspPor3 and ICEValSpa1 具备有活性的限制性修饰系统,它们阻止外源 DNA 的入侵。
Bordeleau 等在霍乱弧菌中发现有 3 个 SXT/R391 包含编码二鸟苷酸环化酶的基因,二鸟苷酸环化酶催化 c-di-GMP 的生成,c-di-GMP 调节生物膜的形成、运动能力以及毒力。但是迄今为止,还没有在SXT / R391 中发现直接的毒力基因。 Rodríguez-Blanco 等在 Enterovibrionigricans 中首次发现 SXT/R391 包含对季胺盐类化学消毒剂具有耐受性的基因 qac,扩展了对 SXT/R391 功能的认识。当然,我们也不排除可变区的一些基因为潜在的抗生素耐药性提供编码,因为抗生素的使用历史仅几十年,而耐药基因的出现和进化不可能在几十年内完成。目前已知的 SXT 多数包含耐药基因 strB、sull、floR、dfrA,分别提供链霉素、磺胺甲基异恶唑、氯霉素、甲氧苄啶抗性,R391 包含卡那霉素耐药基因。但在SXT / R391 中也逐渐发现了一些新的耐药基因,例如: 头孢菌素耐药基因 blaCMY-2和利福平耐药基因。SXT/R391 的大多数耐药基因位于复合转座子或整合子的基因盒中。转座子和整合子种类多样、携带的耐药基因高度多样化,因此我们相信未来还会在 SXT/R391 中发现更多的耐药基因。
3、 SXT /R391 调控及环境因子对其转移的影响
在整个 SXT/R391 调控中,SetC/SetD 是最关键的转录激活蛋白,研究表明至少它们是 int 以及系列tra 基因转录的激活蛋白,setC/setD 的转录则受到一种与 λ 噬菌体阻遏蛋白 cI 高度相似的 SetR 阻遏蛋白的抑制。Beader 等发现: 霍乱弧菌MO10 的 SXT 中 setCD、setR 分属于不同的操纵子,二者的启动子( PL、PR) 位于 s086( setCD 上游,同属1个操纵子) 与 setR 基因之间,SetR 可与和 s086 间 3个位点结合( 操纵序列) 控制转录活性。SetR 也与自身启动子 PR下游操纵序列结合,表明 setR 的表达是自我抑制的,setR 基因无 SD 序列。setCD、setR 操纵子的这些结构和调控特征完全类似于 λ 噬菌体溶原/非溶原开关系统的结构与调控特征。
SetCD 是 1 个很强的激活蛋白,不仅激活 SXT 相关基因的转录,对一些带有 int 基因的 MGE,SetCD 也可以激活其整合酶基因转录,从而促进转移,例如:SetCD 的激活也驱动宿主基因岛的转移,这主要是SetCD 对基因岛 int 及 rdfM 的激活所致。SXT / R391 的转移受环境因子影响明显。一些可以引起 DNA 损伤的环境因子( 如: 丝裂霉素 C、紫外照射、环丙沙星) 均可明显增加 SXT/R391 的转移频率。这些 DNA 损伤环境因子可引起细菌的SOS 反应,进而激活 RecA 活性,类似于 RecA 对LexA 阻遏物的水解,RecA 对 SXT / R391 的 setCD 阻遏蛋白 SetR 也产生水解作用,进而激活了 setCD 表达,SXT/R391 得以高频转移。
4、 SXT /R391 接合转移过程
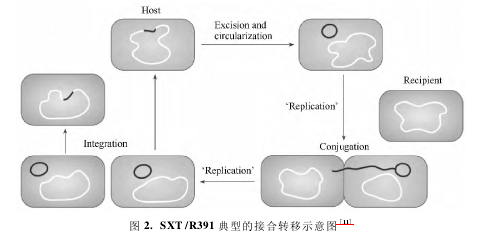
SXT /R391 的接合转移过程如图 2 所示。在 Int和 Xis 的作用下,SXT/R391 从供体菌的染色体 prfC位点剪切,形成线性的 SXT/R391 分子,它的二端分别是具有正向重复的 attL 和 attR 位点,因此二端可以重组,形成新的 attP 位点,线性的 SXT/R391 分子完成环化。oriT 在 1 个解旋酶 TraI 的作用下形成 1个切口,从这个切口开始解链成一个单链 DNA,该单链 DNA 可以通过供体菌和受体菌接合形成的接合通道( 系统) 进入受体菌细胞。SXT/R391等 ICE 也为接合系统编码,推测几乎所有的 ICE 都采用该系统进行 ICE 的转移,该接合系统类似于 IV型分泌系统。IV 型分泌系统组成 1 个蛋白复合物,包含了细胞外的成分,例如: 鞭毛。IV 型分泌系统形成一个跨膜的分泌通道,用于蛋白转运或者与环境中的 DNA 交流。进入受体菌的单链DNA 在受体菌 DNA 聚合酶的作用下再次组装成双链的环形分子,该双链 DNA 分子可以在 Int 的作用下整合入受体菌的特异位点( 通常是在 prfC 基因前) ,此时环形分子的 attP 位点与受体菌染色体的attB 位点也完成重组,产生新的染色体 attB 位点。此外,在原供体菌中,残留的单链的 DNA 再次形成双链,然后重新整合入供体菌的染色体,从而完成 SXT/R391 接合转移过程,在这个过程中 SXT/ R391 并不会因为转移而从原供体菌丢失。
5、 SXT /R391 在宿主系统的稳定性及重组
SXT / R391 并不会轻易从宿主染色体丢失,大多数的 SXT/R391 的热点区 HS2 都包含 mosA 和mosT 基因,它们为毒素-抗毒素系统编码,研究表明包含毒素-抗毒素系统的 SXT 通过杀死或者抑制丢失 SXT 的细胞而维持其存在。一些不存在mosA 和 mosT 基因的 SXT / R391,可能会编码其它类似的 基因来维持其稳定性例如在 R391 和ICEVchMex1( STX 的一种) 中都包含 2 个类似功能的基因( orf1 和 orf3) ,它们 1 个编码 HipA 样的毒素,另1 个编码与抗毒素 HipB 亲缘关系较远的转录抑制子。
SXT / R391 等 ICE 可以阻止细胞再次获得密切相关的同类 ICE,称之为进入排斥。SXT/R391可以划分为 2 种排斥类群,分别为 S 群和 R 群,拥有 S 群的宿主会阻止再次获得另一个属于 S 群的ICE,但不阻止对 R 群 ICE 的获得,反过来也是一样,Eex 对相同类群拷贝的排斥起主导作用,内膜配对形成蛋白 TraG 参与其中。不同类群的不排斥性造成宿主中的 ICE 可以形成串联排列。
实质上,当一个细胞同时进入 SXT 和 R391 型ICE,并形成串联排列时,二者并不能稳定存在,而是发生重组形成杂合 ICE。后来发现,SXT/R391不仅有整合、剪切、转移系统,还自带类似 λRed 重组系统。bet( s065) 、exo( s066) 是这个系统的 2个关键基因,bet 产物 Exo 是核酸外切酶,结合在双链 DNA 末端,降解 DNA 使之产生 3’突击端; exo 产物 Bet 蛋白结合在单链 DNA 上,介导互补单链 DNA退火。与 λRed 重组系统不同,SXT/R391 的重组系统没有基因 gam。然而 SXT/R39 的重组系统更为复杂,bet、exo 位于 1 个大的操纵子之中,这个操纵子还包含 12 个其它的保守基因,其启动子位于基因 s063、s089 之间。bet 上游有 1 个 ssb 基因,主要抑制 Bet/Exo 活性,此外 bet 上游还有 1 个衰减子,抑制重组系统转录。如前所述,引起 DNA 损伤的物理、化学因子可以造成 SXT/R391 转移频率的明显增加,实验还发现引起 DNA 损伤的丝裂霉素C 也可以引起 bet、exo 表达量的明显增加,进而促进重组的发生,与 SXT/R391 转移的诱导不同,DNA损伤因子对 bet、exo 的诱导是非 RecA 依赖的。这从侧面反映了 bet、exo 操纵子调节的复杂性。
SXT / R391 的重组系统促进了杂合 ICE 的形成,SXT / R391 杂合赋予了宿主新的表型特征( 如: 新的耐药性状) ,增加了 SXT/R391 的多样性,因此对于SXT / R391 的进化具有重要意义。
6、 SXT /R391 的广泛转移及分子机制
宿主范围是衡量 1 个传播耐药基因的 MGE 影响力的关键因子。在实验室条件下,SXT/R391显示了相当高的转移频率和宽广宿主范围。例如:
O139 霍乱弧菌株的 SXT 可以转移到大肠杆菌( Escherichia coli) 中,转移频率约为 10- 6/ 受体菌; 美人鱼发光杆菌( Photobacteriumdamsela) 的ICEPdaSpa1 不仅可以转移到发光杆菌属的细菌中,也可以转移到大肠杆菌中; 在奇异变形杆菌中发现的 SXT/R391 家族的 ICEPmiJpn1,能够转移到大肠杆菌、肺炎克雷伯氏杆菌( Klebsiellapneumoniae) 、肠沙门氏杆菌 ( Salmonella enterica ) 以 及Cirobacterkoseri 中,从而赋予这几种受体菌对头孢菌素的耐药性; 灿烂弧菌 ( Vibrio splendidus) 、Enterovibrionigricans 的 SXT / R391 也可以转移到大肠杆菌中,并赋予其相应的耐药表型,转移频率介于10- 5- 10- 6/ 受体菌。从中可以看出,虽然供体菌和受体菌的亲缘关系很远,但仍然具有相当高的转移频率。我们推测 SXT/R391 具有广泛的宿主范围主要由 3 种原因引起: 第一,决定 SXT/R391 是否转移的关键因素在于受体菌是否具有与其 attP 位点相似的 attB 位点,当存在 attB 位点时就可以发生转移,转移与细菌的种属没有明显关系。根据 SXT/R391 对 attB 位点的识别特征,加拿大 Burrus 实验室研究人员已经在多个载体或菌株中人为引入 attB位点,用于定点捕获 SXT/R391( 未出版) ; 第二,attB位点并非要求完全与 attP 相同,研究显示 SXT/R391 attP 位点与受体菌 attB 位点可以允许高达23. 5% 的错配; 第三,我们通过对已知的 attL 和attR 排列后发现,不同 SXT / R391 的 attL( 或 attR) 长度不超过 17bp( 未出版) ,较短的 attL、attR( SXT/R391 环化后形成 attB) 无疑加大了 SXT / R391 匹配到整合位点机率。由于 SXT/R391 具有高的转移频率、潜在的广泛宿主范围,我们推测 SXT/R391 具有迅速传播并扩大分布区域的趋势。
7、 SXT /R391 的扩散趋势及潜在危害
如前所述,目前已经有超过 40 个 SXT/R391 被发现,包括普罗维登斯菌、发光杆菌、施万氏菌、变形杆菌以及弧菌属的多个物种,以弧菌中发现最多。
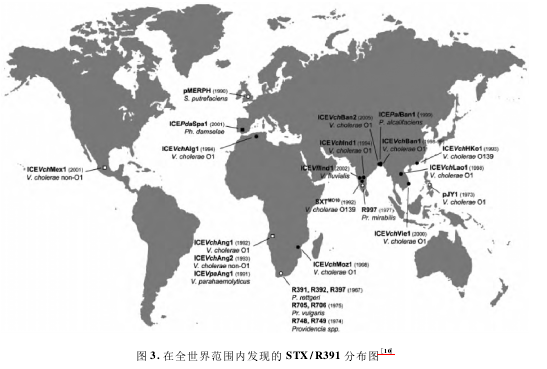
2006 年,Burrus 等根据已经报道的携带 SXT/R391 的细菌株分离地点,描绘了 SXT / R391 的世界分布图( 图 3) 。从图中可以看出,已知的 SXT/R391主要出现在非洲和亚洲,而美洲、欧洲、大洋洲则几乎没有报道; 无论临床株还是环境分离株,所有的SXT / R391 均出现在沿海地带,这一结果结合 SXT /R391 的来源菌多为海洋细菌,显然说明海洋环境很可能是 SXT/R391 发源地,也是其目前主要的贮藏库,并且经由海洋环境株向临床株扩散。目前,SXT / R391 主要出现在非洲和亚洲,并没有呈现出世界性分布,结合 SXT/R391 的转移特征,我们推测SXT / R391 很可能仍然处在一个扩大分布范围的过程中。2012 年,Rodríguez-Blanco 等发现: 2002 至2010 年分离 自西班牙的溶藻弧菌( Vibrioalginolyticus ) 、 灿 烂 弧 菌、 Vibrio scophihalmi、Shewanellahaliotis、Enterovibrionigricans 的 498 株细菌中有 12 株携带 SXT/R391( 2. 4%) ; 令人感兴趣的是 2009 -2010 年分离的菌株虽然只占到总菌株数的 16.6%,但是却有一半的 SXT/R391来源其中,这一结果表明日益增加的选择压力很可能加速了 SXT/R391 的传播。2004 年,Beaber 等在 Nature 发表文章指出: 1993 年以前从未在霍乱弧菌中发现 SXT 元件,但是仅仅 10 年时间,几乎所有的临床霍乱弧菌株都携带有 SXT/R391,这表明SXT / R391 的水平转移相当迅速; Beaber 等同时证实: 无论霍乱弧菌或 SXT 受体菌大肠杆菌都会在SOS 反应的刺激下明显增加 SXT 的转移频率。例如: 丝裂霉素 C 和喹诺酮类药物环丙沙星的使用可以造成 SXT 的转移频率增加几百倍以上。因此Beaber 等指出: 喹诺酮类抗生素( 如: 环丙沙星)的广泛使用可能会促使 SXT 及相关 ICE 的广泛传播。Beaber 等的报道警示我们应当谨慎对待喹诺酮类以及其它抗生素药物的使用问题。同时我们也不排除其它的一些引起细菌 DNA 损伤的物理、化学因子可以促进 ICE 的传播。
喹诺酮类药物是近 30 年才被广泛使用的抗生素,属当前主流的抗生素种类。结合 Rodríguez-Blanco 等、Beaber 等的研究结果以及 SXT/R391 分布特征分析,我们有理由推测喹诺酮类等抗生素的广泛使用,很有可能造成携带 SXT/R391 的细菌在临床和环境中出现的频率逐渐增高,并且借助海洋水流以及人口流动,含 SXT/R391 的细菌株势必会在全球扩散。2012 年,我们对2006 - 2009 年从华南沿海分离的 192 株溶藻弧菌携带 SXT/R391 的情况进行了调查,结果发现 17 个株 SXT/R391 检测阳性,出现比例达 8. 9%,种系发生分析表明溶藻弧菌 SXT/R391 部分序列与 5 个细菌属的 SXT/R391 高度相似,这 5 个属分别是弧菌属、普罗威登斯菌属、变形杆菌属、发光杆菌属、施万氏菌属。本结果表明华南沿海分离的溶藻弧菌SXT / R391 不仅出现比例高,而且元件本身的多样性丰富,这是第一次报道溶藻弧菌中存在 SXT/R391。Rodríguez-Blanco 等的报道中也有 1 株溶藻弧菌携带 SXT,但 SXT/R391 出现率明显低于我们的报道。2013 年笔者对加拿大东西海岸分离的总计 289 株溶藻弧菌、副溶血弧菌、创伤弧菌( 分离于 2008 - 2011 年) SXT/R391 携带情况进行调查,没有发现其携带 SXT/R391( 未出版) 。在华南沿海分离溶藻弧菌中 SXT/R391 的高出现率表明SXT / R391 很可能已经在中国以及周边国家的海洋中盛行。值得注意的是: SXT/R391 不仅能够实现自我转移,还可以通过共整合的方式,驱动与之没有任何遗传关联的质粒和染色体基因岛的转移。质粒不仅可以携带耐药基因,也可以携带毒力基因; 一些重要细菌病原的关键毒力因子也被发现位于基因岛上,如: 霍乱弧菌、副溶血弧菌、大肠杆菌等,因此 SXT/R391 也可能促进毒力因子的扩散。鉴于 SXT/R391 等 ICE 转移和盛行带来的风险,卫生部门以及医学微生物科学家应对其扩散保持充分警惕。
8、 问题和展望
在抗生素药物使用的 70 余年里,人类已经向地球生物圈输入了大量的抗生素以及其它化学药物,它们已经显著改变了微生物赖以生存的环境,破坏了微生物间长期天然存在的拮抗平衡,微生物也通过自身的调整来适应这种变化。通过一系列机制捕获耐药以及其它利益化的基因是细菌对环境做出适应性的表现,同时也是细菌进化的一种重要驱动力。因此,抗生素药物的广泛使用、滥用及不合理处置是携带耐药基因的可移动元件广泛传播,新耐药菌、超级细菌频现的幕后推手。抗生素类药物在人类被滥用的情况可能还不是最严重的问题,养殖业才是抗生素药物滥用的重灾区,除了少数发达国家,养殖业中抗生素药物的滥用更为普遍,而且养殖废水、废物的排放也没有严格的管制,这种情况在中国尤为严重。这些现状可能是耐药基因传播、环境细菌广泛产生耐药性的问题根源。绝大多数的 SXT/R391 携带株出现在亚洲和非洲地区似乎与这些地区过度使用抗生素以及比较随意的排放含抗生素污水存在联系,这也是一个值得探索的问题。
SXT /R391 可以携带多种耐药基因以及其它未知功能的基因,并且具有很强的自我转移频率,SXT / R391 潜在的宿主范围宽广。SXT / R391 具有多个外源 DNA 插入位点,外源 DNA 形成的可变区在每一个元件中的总长度可以达到 30 -60 kb,比起质粒,SXT/R391 显然具有更强的容纳外源 DNA 的能力,目前已知的 SXT/R391 通常携带 4 种耐药基因,但是我们推测在环境选择压力下,SXT/R391 完全可能捕获更多的耐药基因以及其它可以赋予宿主抗逆功能的基因,如果没有充分的认识以及必要防范措施,以 SXT/R391 为载体的更多新型耐药菌株甚至超级细菌的频现将会成为趋势。虽然我们搞清楚了 SXT/R391 的基本结构以及部分基因的功能,但是对于它仍然知之甚少,因此还需要持续不断的探索。





