摘 要: 20世纪90年代以来,随着国家、社会对水环境质量的日益关注,水生植物这一小众的生态类群在我国受到了越来越多的重视。从水生植物多样性编目、面临的危胁(湿地退化、水体富营养化、全球气候变暖、外来物种入侵)、相关的基础性研究(遗传多样性、土壤种子库、种子活力)和保护实践(自然保护区、种质资源库)等方面,对20世纪90年代以来的水生植物多样性研究工作进行梳理与展望,以期为后续更好的保护提供建议。
关键词: 水生植物; 生物多样性; 综述;
Abstract: Since 1990 s, with the increasing attention to the quality of water environment, aquatic plants as a small ecological group has received more and more attention in China. Research on the diversity of aquatic plants since 1990 s are concluded from the aspects of catalogue of aquatic plant diversity, threats of aquatic plant diversity(wetland degradation, water eutrophication, global warming, invasion of alien species), basic researches(genetic diversity, seed bank, seed vitality maintenance mechanism) and conservation practice(nature reserve, germplasm bank), with a view to providing recommendations for better follow-up protection.
Keyword: aquatic plant; biodiversity; summary;
1、 水生植物多样性编目
1.1、 定义与范围
生物物种名录是一个自然地理区域或行政单元的生物区系本底性的基础资料,及时更新生物物种名录对于生物多样性研究和保护与利用实践都是十分重要的。水生植物编目面临的第一个问题就是哪些植物是水生植物。实际上至今为止仍然没有一个普遍被人接受的水生植物定义,李伟等对不同的水生植物定义进行了比较细致的分析[1,2]。
以国内经常使用的水生植物专着为例,《中国水生高等植物图说》[3]收录水生高等植物54科、120属、295种。《中国水生维管束植物图谱》[4]收录了水生维管束植物317种,共有61科、145属。最近出版的《中国水生植物》[5]则记载了741种水生植物。再比较诸如《中国湿地植被》[6]等提供的水生植物名录,可以发现中国水生植物的种类记录从数百种到上千种不等。这一差别的基本原因就是不同研究者对水生植物的定义存在不同所致。如果不考虑各研究中的水生植物定义,在开展比较分析时往往会得出荒谬的结论。在无法就水生植物定义取得一致的情况下,做比较研究时采用狭义的水生植物定义是妥当的[2]。
水生植物编目工作中面临的另一个常见问题就是我国有多少种水生植物。在此按照国际上有关水生植物的一般认识,对我国水生植物的种类数目做个基本估算。
所有的水生高等植物都是从陆生祖先进化而来的,而陆生植物又是从水生绿藻很可能是双星藻纲(Zygnematophyceae)进化而来[7],因此包含了能够适应空气和水中生活的进化痕迹。约17%的现存被子植物科包含水生植物,大概有6 000多种[8,9],不足所有被子植物总数的2%[10]。以2%作为水生植物种类的估算比例,我国水生植物的种类数目大致在600种左右(《中国植物志》记录了31 142种植物)。
在实际操作中建议采用以下方法:① 按照Cook的建议,水生植物限定于草本植物,并且Cook给出了一个比较完整的包含水生植物的科属名录[8],可以作为确认水生植物的参考;② 考虑对象种在全部/主要分布区内的种群所在生境条件,水生植物是以湿地(按Ramsar公约定义)为生境的植物。

水生植物一般被分为挺水植物、浮叶植物、沉水植物和漂浮植物等4类。而在实际应用中往往又提出湿地植物或者湿生植物与之并列,在这些工作中,一般把湿地(湿生)植物作为相较挺水植物更耐旱的一类植物。实际上前4类的划分主要依据的是植物的光合器官与水位和基质的关系,而湿地(湿生)植物从语义上看是能够生长于湿地生境中的植物,与前4类的划分基础不一致。为避免混乱,可以认为湿地(湿生)植物等同于挺水植物。如果需要在这两类植物间作出区分,从生理过程考虑有可能提供一个相对科学的划分。从目前湿地(湿生)植物大多数应用场景看,这些植物具备了从相对中生到旱生生境的适应能力,因此可以采用以下标准对两类植物进行区分:在生长于湿地生境的植物中,主要种群在生活史过程中普遍出现耐旱生理过程甚至耐旱结构的植物可以被称为湿生植物,不出现耐旱生理过程或者耐旱生理过程不能支持植物完成正常生活史过程的植物可归类于挺水植物。
1.2、 水生植物多样性编目成就
中国的生物分类学者艰苦奋斗百余年, 基本摸清了中国生物物种的家底。特别是近60多年来,《中国植物志》《中国动物志》和《中国孢子植物志》的编研,极大地提高了中国生物区系的认知水平和生物分类学的发展速度[11]。
我国水生植物多样性保护的基础性工作也主要以编目为主,尤其是“三志”的编研为水生植物多样性编目工作的开展起到了积极的推动作用,在各类志书编撰过程中积累的大量文献、标本为水生植物多样性编目工作的开展提供了大量第一手资料。其中《中国植物志》第八卷[12]的编研过程极大地促进了我国水生植物在系统学、分类学、生态学等方面的研究,为我国水生植物多样性保护提供了丰富的种类、分布、生境、标本等方面的资料。
从中国数字植物标本馆(http://www.cvh.ac.cn)提供的信息看,大量的水生植物标本几乎都与《中国植物志》的编撰有关。在这期间出版的《中国水生高等植物图说》《中国水生维管束植物图谱》至今仍然是水生植物研究工作中最常用的文献资料。
总体上看,21世纪以前我国水生植物的编目工作主要依赖于“三志”编研中积累的大量资料,除了《中国植物志》第八卷外,水生植物的调查通常依附于其他类群的相关考察工作中。因此,尽管水生植物只是一个种类数目有限的生态类群,而且《中国植物志》的编撰工作已经告一段落,随着调查研究工作的持续开展,仍然有水生植物的新分类群在不断发现。例如在广西桂林及百色等地发现了灌阳水车前(Ottelia guanyangensis)和凤山水车前(O. fengshanensis)两个海菜花属新种,分子学证据证明了这两个“好种”的成立[13,14]。
进入21世纪以来,随着国家、社会对水环境问题的日益重视,在《国家中长期科学和技术发展规划纲要(2006-2020年)》《国家基础研究发展“十二五”规划》《全国生物物种资源保护与利用规划纲要》等指导下,针对水生植物(2013~2018年)、入侵植物(2015~2018年)、沼泽植物(2020年起)等类群均启动了科技基础资源调查专项,这些项目的完成能够为我国水生植物多样性现状提供更翔实的现状数据。
2、 水生植物多样性面临的威胁
2.1、 湿地退化对水生植物的威胁
21 世纪以来,受经济发展、城市化过程、气候变化影响,湿地退化已成为一种全球现象[15]。在我国,湿地退化现象严重,重要标志之一即是湿地面积的急剧下降[16],据《中国湿地保护行动计划》[17]统计,全国围垦湖泊面积达130万hm2 以上,超过了我国现今五大淡水湖面积之和,由于湖泊围垦而失去的调蓄容积在350亿m3 以上,因围垦而消亡的天然湖泊近1 000个。根据环境保护部《中国履行〈生物多样性公约〉第四次国家报告》[18]:我国约有40% 的重要湿地面临严重退化的威胁。随着湿地退化,洪涝渍害加重,湿地水生植被种类减少, 湿地生物多样性明显下降,优势物种分布区缩小,群落结构发生改变,珍稀濒危物种减少或灭绝,局部区域甚至出现生态环境恶化。
2.2、 水体富营养化对水生植物的威胁
世界范围内营养物质大量输入所引起的水体富营养化是全球环境剧变的表现形式之一[19,20]。我国湖泊富营养化问题越来越严重,研究表明,到2010年为止,我国长江中下游、东北、云贵高原大于10 km2的138个湖泊中有85.4%的湖泊出现不同程度的富营养化[21]。伴随水体富营养化的加剧,水生植物的种类、数量显着降低;水生植物由以清水生长为主的植物类群逐渐变化到以耐污植物为主的植物类群[22]。以云贵高原湖泊面积大于10 km2湖泊为例,20世纪80年代初,每个湖泊至少含有6大不同类型水生植物群落[23],而近年来的调查显示,有近半数的湖泊水生植被完全消失。植物群落结构主要从以海菜花(Ottelia acuminata)等为主的植物类群转变为篦齿眼子菜(Potamogeton pectinatus)、狐尾藻(Myriophyllum verticillatum)等耐污种为优势的群落类型[24]。
2.3、 全球气候变暖对水生植物的威胁
全球气候变化导致生物多样性降低进而引起的不同生态系统共同面临的重大科学问题[19,20,25,26,27,28]。IPCC第四次报告[29]指出,近百年来全球平均地面温度已上升0.4℃~ 0.8℃,达到1 000 a以来的最高值。气候变暖加速了全球的水文循环,降水量增加,洪水和干旱等极端事件增多,逐渐改变了水生植被赖以生存的水环境格局,导致了湿地的退化以及外来物种的入侵[29,30],严重威胁水生植被的生存和群落维持。同时,气候的变暖直接导致高山区域超过40%的冰川正在或已经消失,过去的50 a间,我国西北冰川面积减少了21%[31,32]。冰川的融化导致高原沼泽、湖泊理化性质的改变,破坏植物多样性和生态系统的平衡。
以往全球气候变化对植物影响的研究主要针对陆地生态系统开展[33,34,35],对水生植物影响的工作非常零星。最近Iversen et al. 在Science上撰文指出:在全球范围内,具有HCO-3利用能力的水生植物出现的频率随着生境HCO-3浓度的增加而增加[36];在区域尺度上,当水域生境中CO2浓度大大高于CO2的空气-水平衡浓度的地方,具有HCO-3利用能力的水生植物种类出现的频率就降低,这与HCO-3利用能力是水生植物对生境碳限制的一种适应的结论一致,而全球气候变化所带来的HCO-3和CO2浓度的变化可能会改变淡水植物群落的物种组成。由于水生植物所在生境在水分条件与无机碳环境方面与陆生植物完全不同,全球气候变化对水生植物影响的研究有可能在水生植物的适应性进化方面取得突破性的成果。
2.4、 外来物种入侵对水生植物的威胁
外来物种入侵会对入侵地的生态系统产生严重的破坏,是群落生物组成、结构和生态系统功能衰退的原因之一[37,38,39]。外来种入侵对本地生物多样性也具有毁灭性的影响[36],在威胁生物多样性的原因中,其被认为是仅次于生境破坏的第二大威胁。全球气候变化以及日益频繁的人类活动则对生物入侵的发生起到了促进作用 [40]。与其他类群相比,水生植物由于特殊的生活史特征与生态适应性,使得水生植物类群中外来种类的比例远远高出其他类群,并且水域生境的连通性导致水生植物入侵的危害常常具有突发性与全局性,更难以预测与控制。植物群落对入侵物种的抗性与群落的物种多样性有密切关系,一般认为多样性程度高的群落比多样性低的群落能更有效地抵御外来种的入侵,多样性的损失是导致入侵发生的一个重要原因[41,42,43]。我国目前水生入侵植物的快速扩散,一方面与生境破坏带来的本土水生植物多样性丧失有关;另一方面也与外来水生植物的人为扩散有着紧密关系。近一二十年来,随着全社会对水环境质量的日益重视,退化水域生态系统的恢复重建已经成为全社会的共识,在几乎所有的退化水域生态系统恢复重建工作中,都将水生植被的恢复重建作为一个重要目标与手段。然而在实际操作过程中,大量的外来水生植物堂而皇之地进入到生态系统恢复重建工程中,这对本土水生植物多样性的保护不能不说是一种讽刺。
3、 水生植物多样性保护的基础性研究
水生植物由于特殊的进化历史与生境条件,在生活史中表现不同于陆生植物的生活史特征[44],水生植物的保护生物学研究依然是指导其多样性保护的重要基础和指导。在此,主要从遗传多样性、土壤种子库和种子活力3方面对相关基础性研究进行总结。
3.1、 水生植物的遗传多样性研究
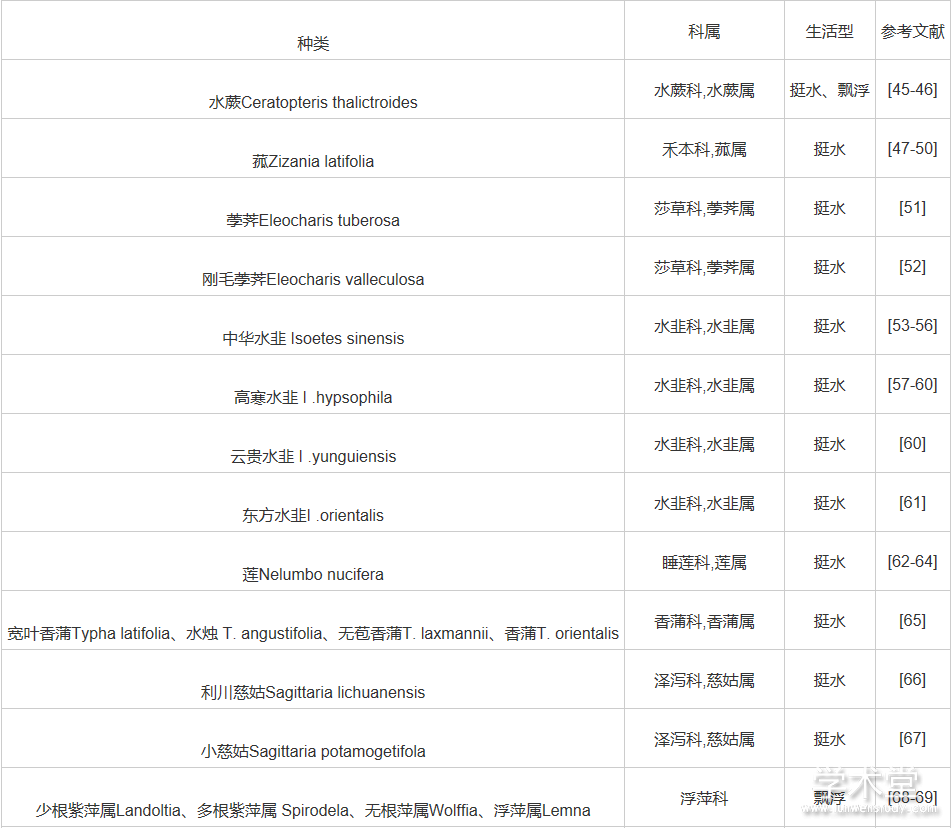
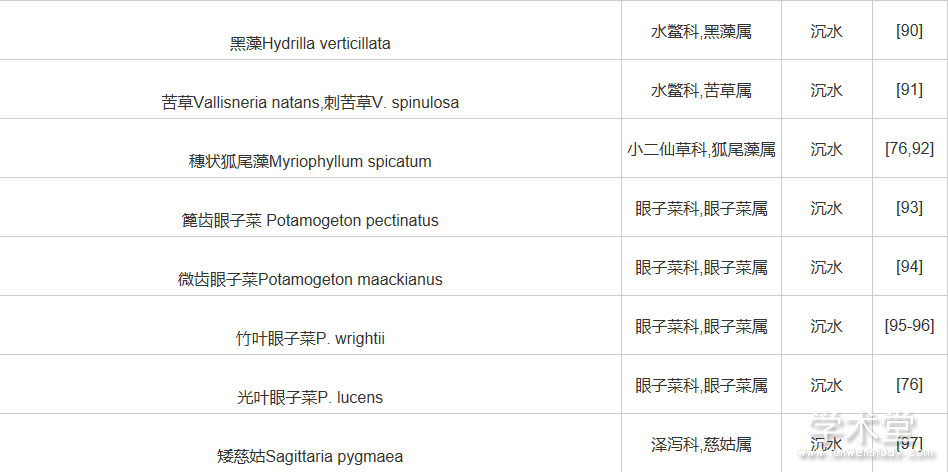
随着植物分子生物学研究的不断发展,利用各种分子标记技术开展水生植物遗传多样性分析评价的工作也随之迅速发展。不过总体上看,与陆生植物相比较,我国水生植物遗传多样性的研究起步晚,并且研究基础较为薄弱。表1对目前已经开展的水生植物遗传多样性研究进行了总结。
表1表明遗传多样性研究主要在眼子菜科、水鳖科、泽泻科等水生植物代表性类群中开展。即使覆盖的分类群尚需扩展,现有结果仍然对水生植物的多样性保护提供了宝贵的指导性意见:① 尽管营养繁殖是水生植物主要的繁殖方式,但是主要水生植物并未表现出较低的遗传多样性;② 尽管水生植物由于所处水环境的连通性较高,很多水生植物表现出较为广泛分布的特点,但是水生植物不同种群间仍然表现出明显的遗传分化。
3.2、 土壤种子库研究
种子库是指土壤表面或基质中具有繁殖能力的种子、果实、无性繁殖体以及其它能再生的植物结构的总称[98]。狭义的种子库通常专指种子和包含种子的果实。
湿地一直是种子库研究的重点生境,很多种子库研究工作中的重要结论均是在湿地研究中取得的。我国湿地种子库的研究起步很晚,从2000年起才逐步开展[99,100]。湿地种子库在湿地的保护和受损湿地的恢复与重建中具有重要作用:充分利用原湿地保留的种子库,以及通过种子库移植等方法恢复湿地植被,将更有利于湿地的物种多样性和遗传多样性的恢复[101]。
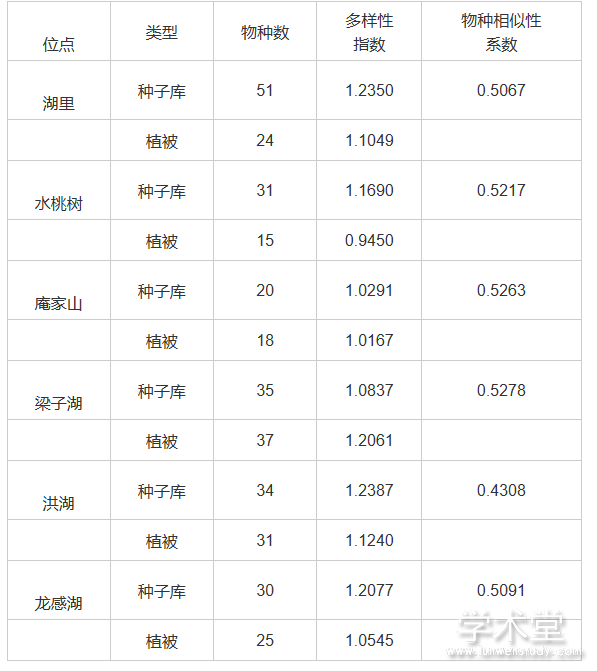
以长江中下游不同湿地种子库研究为例,种子库中物种数远高于同期的地表植被(见表2),除梁子湖外, 其他5个沼泽的种子库中含有比地表植被中更多的物种。在湖里沼泽和水桃树沼泽中, 种子库的物种数是地表植被的两倍。种子库的Shannon 多样性指数通常高于地表植被[102]。
表1 我国水生植物遗传多样性研究工作总结

湿地土壤种子库不仅常常包含比地表植被丰富的植物种类,而且还常常表现出与地表植被不同的遗传多样性,因为土壤种子库通常累积了不同时期和环境条件下地表植物产生的种子。由于湿地的地表植物种群具有更高的灭绝风险,因此当地表植被受到突发的干扰破坏后,种子库具有恢复种群遗传多样性的潜力[102,103]。种子库作为遗传基因库,可以用于种群恢复,在珍稀濒危植物的原位保护中具有重要的作用。
表2 长江中下游湿地种子库与地表植被物种多样性比较
3.3 、水生植物种子活力维持机制
Pitman 和 Jorgensen[104]统计表明,全世界约有50%的植物物种受到灭绝的威胁,如不及时采取有效的保护措施,到21世纪末将有2/3的维管植物消失。迁地保护已经成为保护植物生物多样性的重要措施之一,其中建立以收集种子为主体的基因库是当今保护植物种质资源最为普遍、且可靠易行的方式,在世界范围内库存的约6 100 000份种质资源中,近90%是以种子形式保存的。种子保存是植物种质遗传多样性迁地保护的一种廉价而有效的方法。
利用种子进行植物的迁地保护,最为重要的就是保护种子的活力。如何有效地延长种子的贮藏寿命和最大程度地减少种子在贮藏过程中细胞的衰老损伤,对于植物种质资源的保存至关重要。陆生植物种子保存已经进行了广泛而系统的研究,低温、干燥贮藏仍是目前基因库中种子种质保存的主要方法。然而干燥贮藏只适用于正常型种子(orthodox seeds),对于顽拗型种子(recalcitrant seeds)的贮藏则必须采用另外的方式。
与陆生植物相比,国内外对水生植物种子生理的研究非常之少,主要工作集中在一些种类的种子萌发方面,有关水生植物种子贮藏方面的基础数据非常缺乏,这给利用种子进行水生植物迁地保护带来了困难[105]。目前已知具顽拗型种子的水生植物种类有凤眼蓝(Eichhornia crassipes)、菱(Trapa natans)、菰(Zizania latifolia)、大米草(Spartina anglica)和南亚野稻(Porteresia coarctata)等[106]。而绝大部分水生植物的贮藏类型还是未知的,其有效的贮藏方法更有待于作进一步的研究。
虽然水生植物的种子一般被认为是顽拗型种子,不宜采用目前陆生植物种子保藏的通用方法,但是目前的野外调查发现,水生植物的种子在野外黑暗、厌氧、饱和水分条件下可以长时间地保持活力:对洪湖湖滨湿地种子库的研究表明,在地表下25 cm处仍然可以发现相当数量具有活力的苦草和黑藻种子[107],如果仅从洪湖的沉积速率来看,这些深度埋藏的种子的寿命是相当长的。在其他一些湿地种子库研究工作中,同样可以在埋藏较深的土层中发现相当数量的水鳖科植物种子[108]。
研究表明,当年成熟的水鳖种子在干燥条件下贮藏,其活力会急剧降低,表现出典型的顽拗型种子特征。但是埋藏于水底泥沙中的水鳖种子在两三年后依然保持非常高的活力[105]。这些现象和研究工作表明,水生植物在适应水环境的进化过程中,有可能发展出了特殊的种子活力维持机制。对这些机制开展系统深入的研究有可能为利用种子保藏技术来保护水生植物多样性提供技术和理论指导。
4 、水生植物多样性保护实践
4.1、 自然保护区
自然保护区在全球范围内的广泛设立,是当代自然资源保护和管理中的一件大事。截止到2018年5月,我国共建立国家级自然保护区474处。此外,各省市也根据实际条件建立了各级保护区。以湖北省为例,目前有国家级自然保护区22处,已建立省级自然保护区78处,其中与湿地保护相关的省级自然保护区7处。在这些保护区中,除了若干以红树林保护为目标的保护区外,没有任何一处保护区以保护水生植物为目标,但是客观来讲,保护区的建设仍然为水生植物的保护提供了基础。
以龙舌草的调查工作为例,龙舌草是水鳖科海菜花属中分布最广的植物,由于其特殊的光合作用特征近年来受到了高度关注[109,110],热带分布的龙舌草有可能为相关研究工作的深入提供关键的材料。据文献与标本记载,海南是其分布的一个重要区域,不过根据这些信息的调查均以失败告终:这些记录中标明的龙舌草产地已经由于各种人为因素的影响而变得面目全非。但是最近在海南邦溪省级自然保护区这一过去调查中没有记录的地方找到了这种珍贵的植物,尽管这个保护区是为了保护国家一级保护动物海南坡鹿而设立的。
4.2、 种质资源库
迁地保护已经成为保护植物生物多样性的重要措施之一,对于某些可能在自然或人类主导的生态系统中丧失的多样性成分,迁地保护通常通过种质资源圃的作物品系、基因库中的试管苗、保护站中的树种、植物园或种子库(常规和超低温)中的种子等方式对其加以保存,作为备份。
4.2.1、 水生植物活体种植园
目前我国以野生水生植物迁地保护为目标的种质资源圃主要以中国科学院武汉植物园为代表。21世纪以来,武汉植物园在全国开展了大规模的水生植物野外引种和物种保育工作,并在园内建成了水生植物专类园(包括水生植物种质资源圃)和湿地植物专类园。目前武汉植物园“水生植物种质资源圃”保育了包括中华水韭、海菜花、普通野生稻、粗梗水蕨在内的水生维管束植物近500种,建成了我国涵盖量最大、物种最为丰富的水生植物种质资源库。
水生经济植物种质资源圃主要在农业系统的院所建立,其代表是国家种质武汉水生蔬菜资源圃,主要从事水生蔬菜种质资源的征集、保存与评估、遗传育种、脱毒快繁技术、栽培研究及推广应用。目前已保存从全国20多个省份及印度、泰国、马来西亚、新加坡等国家征集的莲藕、茭白、芋、菱、荸荠、慈姑、水芹、芡实、豆瓣菜、蕹菜、莼菜、蒲菜等12类水生蔬菜种质资源1 800余份,另收集保存有睡莲、花菖蒲等水生观赏植物100余份。
4.2.2 、水生植物种子保藏
种子保存是植物种质遗传多样性迁地保护的一种廉价而有效的方法,已得到科学家、政府相关部门和非政府组织越来越多的认可。种子库通常以国际共识的标准建立,以便使种子相对容易收集,能代表物种内的多样性,并能被储藏在相对小的空间里。在设施规模和范畴上,种子库保存的规模都比较大。
2007年2月,国家重大科学工程——中国西南野生生物种质资源库(“种质资源库”)在中国科学院昆明植物研究所初步建成并投入试运行。2009 年11月,种质资源库通过了国家发改委组织的专家验收。从种质资源库建成伊始,水生湿地植物的种子收集保藏就成为其中的一部分工作内容,科技基础性工作专项重点项目:青藏高原特殊生境下野生植物种质资源的调查与保存(2007~2012年)即列出专题任务“青藏高原湿地等植物种质资源调查”。
我国国家重要野生植物种质资源库(National Wild Plant Germplasm Resource Center)以中国西南野生生物种质资源库为核心,于2017年正式成立,依托昆明植物研究所建设和运行,联合全国范围内从事野生植物种质资源收集保藏的相关单位,围绕国家战略需求持续开展重要野生植物种质资源的标准化收集、整理、保藏工作。
至今为止,该资源库中收集保藏的水生植物种子仍然极其稀少,一方面是因为正常情况下,营养繁殖是水生植物的主要繁殖方式,难以收集到足够量的种子用于保存;另一方面也是因为大量水生植物种子的贮藏类型尚无基础数据,无法指导水生植物的种子保藏。
5 、水生植物多样性保护展望
与陆生植物种质资源保护的研究与实践相比,水生植物受到的关注远远落后于其多样性保护的需求。水域湿地是目前最受威胁的生境类型之一,日益频繁剧烈的人为活动所伴随的生境丧失、片段化、污染与富营养化等极大地威胁着水生植物的生存发育,同时全球气候变化导致的水文过程变化进一步改变了水生植物所处生境的水文情势,使得许多水生植物在很短的时期内种群规模极度衰减乃至濒危甚至消失,导致水生植物的灭绝速率远高于陆生植物,其濒危现状更为严重[111]。对水生植物开展迁地保护已经迫在眉睫,这将对以后水生植物的引种回归、种群复壮,以及利用广布种对湖泊进行生态修复都具有重要意义。
5.1、 活体种质资源库
以武汉植物园为例,在其迁地保存的众多水生植物种类中,主要采用采集有限个体进行营养繁殖的方法保存,这种保存方法极有可能造成所保育物种的种质退化[112]。
这种集中式迁地保存方法存在以下难以克服的问题:① 正常情况下水生植物一般依赖其发达的无性繁殖方式来完成种群的更新[64],由于采集条件和栽培空间的限制,往往只能采集有限个体进行迁地栽培,无法保证迁地种群的遗传多样性。② 水生植物迁地保护种群的管理比陆生植物的管理困难。尽管大多数水生植物是多年生的,但是其种群动态类似于陆生一年生植物[64],一般每年都需要从其营养繁殖体而不是种子重新萌发、建群。与陆生植物相比,水生植物的生长发育过程中受同一水环境中其他生物的影响要远远大于陆地生境中其他生物对陆生目标植物的影响,很多水生植物在迁地环境中往往不能长远地保存下去。③ 尽管水生植物一般具有比较广泛的分布区,但是不同的种群在遗传多样性上也常常表现出相当大的变异,为了保存水生植物的多样性,尽可能多地收集不同种群样本必不可少,但是这同样受限于迁地空间。④ 部分分布狭窄的水生植物难以在迁地保护区正常生长。
因此,建议在今后水生植物活体种质资源库的建设中,以分布式代替集中式,即在水生植物的集中分布区,建设各自的活体种质库,统一建设、管理与维护的标准。
5.2、 水生植物种子保藏库
无论是目前的集中式还是今后可能的分布式活体水生植物种质资源圃,所保存的水生植物遗传多样性水平低下的问题仍然存在。真正解决这一问题必须采用陆生植物多样性保存的首要方法——种子库保存。
水生植物特别是沉水植物种子成熟后一般都落在水中,其生境与陆生植物种子很不相同,这可能导致了很多水生植物种子与陆生植物种子有着不同的贮藏和萌发特性。目前有限的研究也发现可以通过一定的贮藏方法来比较长期地保存水生植物的种子活力,表明水生植物尤其是沉水植物种子由于其特殊的生境条件,其贮藏和萌发也有着与陆生植物种子不同的特殊需求。水生植物种子活力维持机制的研究以及相应的水生植物长期贮藏方法的研发,是今后水生植物多样性保护的重要研究课题。
参考文献
[1] 李伟,钟扬.我国内陆水生植被研究概况[J].武汉植物学研究,1991(3):281-288.
[2] 李伟.洪湖水生维管束植物区系研究[J].武汉植物学研究,1997(2):113-122.
[3] 颜素珠.中国水生高等植物图说[M].北京:科学出版社,1983.
[4] 中国科学院武汉植物研究所.中国水生维管束植物图谱[M].武汉:湖北人民出版社,1983.
[5] 陈耀东,马欣堂,杜玉芬,等.中国水生植物[M].郑州:河南科学技术出版社,2012.
[6] 中国湿地植被编辑委员会.中国湿地植被[M].北京:科学出版社,1999.
[7]Wickett N J,Mirarab S,Nguyen N,et al.Phylotranscriptomic analysis of the origin and early diversification of land plants [J].Proceedings of the National Academy of Sciences of the United States of America,2014,111(45):4859-4868.
[8]Cook C D K.Aquatic Plant Book[M].Hague:SPB Academic Publishing,1990.
[9]den Hartog C,Kuo J.Taxonomy and biogeography of seagrasses[A]//Larkum AWD,Orth RJ,Duarte CM.Seagrasses:biology,ecology and conservation[M].Dordrecht:Springer,2006.
[10]Les D,Schneider E.Aquatic origin of monocotyledons[A]// Rudall P J,Cribb P J,Cutler D F,et al.Monocotyledons:Systematics and Evolution[M].Kew:Royal Botanic Gardens,1995.
[11] 马克平.中国生物多样性编目取得重要进展[J].生物多样性,2015,23 (2):137–138.
[12] 孙祥钟.中国植物志第八卷[M].北京:科学出版社,1992.
[13]Li Z,Liao K,Zou C,et al.Ottelia guanyangensis (Hydrocharitaceae),a new species from southwestern China [J].Phytotaxa,2018,361 (3):294–300.
[14]Li Z,Lu M,Gichira W,et al.Genetic diversity and population structure of Ottelia acuminata var.jingxiensis,an endangered endemic aquatic plant from southwest China [J].Aquatic Botany,2019 (152):20-26
[15] Yang Y X.The 21st century hot point and forward position field of international wetland research from Quebec 2000-Millennium Wetland Event [J].Scientia Geographica Sinica,2002 (22):150-155.
[16] Day J W,Yanez Arancibia A,Mitsch W J,et al.Using ecotechnology to address water quality and wetland habitat loss problems in the Mississippi basin:A hierarchical approach[J].Biotechnology Advances 2003(22):135-159.
[17] 国家林业局.中国湿地保护行动计划[Z].2000.
[18] 环境保护部.中国履行〈生物多样性公约〉第四次国家报告[Z].2008.
[19]Bradshaw W E,Holzapfel C M.Climate change,evolutionary response to rapid climate change[J].Science,2006 (312):1477-1478.
[20]Thuiller W.Biodiversity - climate change and the ecologist[J].Nature,2007(448):550-552.
[21] 杨桂山,马荣华,张路,等.中国湖泊现状及面临的重大问题与保护策略[J].湖泊科学,2010,22(6):799-810.
[22]Jeppesen E,Jensen J P,Sondergaard M,et al.Trophic structure,species richness and biodiversity in Danish lakes:changes along a phosphorus gradient [J].Freshwater Biology,2000 (45):201-213.
[23] 李恒.云南高原湖泊水生植被的研究[J].云南植物研究,1980(2):113-139,141.
[24] 于瑶,杜建伟,张汉尧.云南高原湖泊湿地植物群落分布规律研究[J].安徽农业科学,2012,40 (12):7322-7324,7327.
[25] Hughes.Biological consequences of global warming:is the signal already apparent?[J].Trends in Ecology & Evolution,2000,15(2) 56-61.
[26]Sala O E,Chapin F S,Armesto J J,et al.Global biodiversity scenarios for the year 2100 [J].Science,2000 (287):1770-1774.
[27]Montreal D S.Global warming threatens extinction for many species[J].Nature,2000 (497):121.
[28]Pockely P.Climate change transforms island ecosystems [J].Nature,2001 (410):616.
[29]IPCC.Climate Change:Synthesis Report[Z].IPCC Working Group,2007.
[30]Gilvear D J,Mcinnes R J.Wetland hydrological vulnerability and the use of classification procedures:a scottish case study[J].Journal of Environment Management,1994(42):40-414.
[31] Mirza M Q,Vuille M,Bradley R S.Mean Annual Temperature Trends and their Vertical Structure in the Tropical Andes[J].Geophysical Research Letters,2000 (27):3885-3888.
[32]Raper S C B,Braithwaite R J.Low sea level rise projections from mountain glaciers and icecaps under global warming[J].Nature,2006 (439):311-313.
[33]Melillo J M,McGuire A D,Kicklighter D W,et al.Global climate change and terrestrial net primary production[J].Nature,1993(363):234–240.
[34]Cao M,Woodward F I.Dynamic responses of terrestrial ecosystem carbon cycling to global climate change[J].Nature,1998(393):249–252.
[35]Cramer W,Bondeau A,Woodward F I,et al.Global response of terrestrial ecosystem structure and function to CO2 and climate change:results from six dynamic global vegetation models[J].Global Change Biology,2001 (7):357–373.
[36]Iversen L L,Winkel A,Baastrup-Spohr L,et al.Catchment properties and the photosynthetic trait composition of freshwater plant communities [J].Science,2019 (366):878–881.
[37]Diamond J M.Introduction,extinctions,exterminations,and invasions[A]//Diamond JM,Case T J.Community Ecology[M].New York:Harper and Row,1985:65-79.
[38]Wilcove D S,Rothstein D,Dubow J,et al.Quantifying threats to imperiled species in the United States[J].Bioscience,1998(48):607-615.
[39] Mack R N,Simberloff D,Lonsdale W M,et al.Biotic invasions:causes,epidemiology,global consequences and control[J].Ecological Applications,2000(10):689-710.
[40] Dukes J S,Mooney H A.Does global change increase the success of biological invaders?[J].Trends in Ecology and Evolution,1999(14):135-139.
[41]Elton C S.The Ecology of Invasions by Animals and Plants[M].Chicago:The University of Chicago Press,1958.
[42]McGrady-Steed J,Harris P M,Morin P J.Biodiversity regulates ecosystem predictability[J].Nature,1997(390):162-165.
[43] Symstad A J.A test of the effects of functional group richness and composition on grassland invasibility[J].Ecology,2000(81):99-109.
[44] Li W.Environmental opportunities and constraints in the reproduction and dispersal of aquatic plants[J].Aquatic Botany,2014(118):62–71.
[45] 董元火,王青锋.水蕨的生境及其遗传多样性分析[J] .武汉大学学报(理学版),2007,53(2):224-230.
[46]Dong Y H ,Chen J M ,Robert W G,et al.Genetic variation in the endangered aquatic fern Ceratopteris thalictroides (Parkeriaceae) in China:Implications from RAPD and ISSR data for a conservation strategy[J].Bot J Linn Soc,2008,157(4):657-671 .
[47] Chen Y,Chu H,Liu H,et al.Abundant genetic diversity of the wild rice Zizania latifolia in central China revealed by microsatellites[J].Annals of Applied Biology,2012(161):192-201.
[48]Chen Y,Liu Y,Fan X,et al.Landscape-Scale Genetic Structure of Wild Rice Zizania latifolia:The Roles of Rivers,Mountains and Fragmentation[J].Frontiers of Ecology and Evolution,2017( 5):17.
[49] Fan X,Ren X,Liu Y,et al.Genetic structure of wild rice Zizania latifolia and the implications for its management in the Sanjiang Plain,Northeast China [J].Biochemical Systematics and Ecology,2016 (64),81-88.
[50]Zhao Y,Zhong L,Zhou K,et al.Seed characteristic variations and genetic structure of wild Zizania latifolia along a latitudinal gradient in China:implications for neo-domestication as a grain crop[J].AoB Plants,2018,10(6).
[51] 江文,陈丽娟,欧昆鹏,等.荸荠种质资源品种遗传多样性ISSR分析[J].西南农业学报,2015(2):313-318.
[52] 胡波,陈媛媛,李守淳,等.鄱阳湖具刚毛荸荠居群遗传多样性和克隆结构的初步研究[J].武汉植物学研究,2009(27):145-151.
[53] Chen Y,Yang W,Li W,et al.High allozyme diversity and unidirectional linear migration patterns within a population of tetraploid Isoetes sinensis,a rare and endangered pteridophyte [J].Aquatic Botany,2009(90):52-58.
[54] Chen Y,Kong D,Huang C,et al.Microsatellite analysis reveals the genetic structure and gene flow of the aquatic quillwort Isoetes sinensis,a critically endangered species in China[J].Aquatic Botany,2012(96):52-57.
[55] 陈媛媛,叶其刚,黄宏文.中华水韭(Isoetes sinensis)等位酶分析的初步研究[J].武汉植物学研究,2003( 21):91-94.
[56] 陈媛媛,叶其刚,李作洲,等.极濒危植物中华水韭休宁居群的遗传结构[J].生物多样性,2004(12):564-571.
[57] Chen J M ,Liu X,Wang J Y ,et al.Genetic variation with in the endangered quillwort Isoetes hypsophila (Isoetaceae) in China as evidenced by ISSR analysis[J].Aquat Bot,2005,82(2):89-98.
[58] Chen Y,Liao L,Li W,et al.Genetic diversity and population structure of endangered alpine pteridophyte Isoetes hypsophila Hand.-Mazz.revealed by AFLP markers[J].Plant Systematics and Evolution,2010(290):127-139.
[59] Li X,Li S,Chu H,et al.Genetic diversity and population structure of the endangered alpine quillwort Isoetes hypsophila (Isoetaceae) revealed by SSR analysis[J].Biochemical Systematics and Ecology,2013(47):11-20.
[60] 陈进明,刘星,王青锋.中国珍稀濒危特有蕨类植物:云贵水韭的遗传多样性[J].武汉大学学报(理学版),2005 ,51(6):767-770.
[61] 陈进明,王青锋.珍稀濒危蕨类植物东方水韭的遗传多样性[J].武汉植物学研究,2006,24(6):569-573 .
[62]Liu Y,Yang M,Xiang Q,et al.Characterization of microsatellite markers and their application for the assessment of genetic diversity among lotus accessions[J].J.Amer.Soc.Hort.Sci.,2012(137):180-188.
[63] Yang M,Han Y,Xu L,et al.Comparative analysis of genetic diversity of lotus (Nelumbo) using SSR and SRAP markers[J].Scientia Horticulturae,2012(142):185-195.
[64]Chen Y,Wang W,Fan X,et al.Genetic discontinuities and abundant historical gene flow in wild lotus Nelumbo nucifera populations from the Yangtze River[J].Aquatic Botany,2019(158):103-130.
[65]Zhou B,Yu D,Ding Z,et al.Comparison of genetic diversity in four Typhaspecies (Poales,Typhaceae) from China[J].Hydrobiologia,2016,770(1):117-128.
[66]Liu F,Zhao S,Li W,et al.Population genetic structure and phylogeographic patterns in the Chinese endemic species Sagittaria lichuanensis,inferred from cpDNA atpB-rbcL intergenic spacers[J].Botanique,2010,88(10):886-892.
[67] 陈锦华,孙爱珍,汪小凡.小慈姑的遗传多样性和居群分化[J].水生生物学报,2006(5):570-577.
[68] 叶松建,韩冰莹,张家明.福建水生植物浮萍的形态学和分子系统学鉴定及遗传多样性研究[J].热带作物学报,2017(4):101-107.
[69] Xu Y,Ma S,Huang M,et al.Species distribution,genetic diversity and barcoding in the duckweed family (Lemnaceae)[J].Hydrobiologia,2015,743(1):75-87.
[70] Yuan J X,Pan J,Wang B S,et al.Genetic differentiation of Wolffia globosa in China[J].Journal of Systematics And Evolution,2011(49):509–517.
[71] Dong Y H ,Robert W G,Wang Q F.Genetic variation,gene flow and conservation strategy of the endangered aquatic fern Ceratopteris pteridoides in China[J].Ann Bot Fennici,2010,47(1):34-44.
[72] Chen Y,Han Q Cheng Y,et al.Genetic variation and clonal diversity of the endangered aquatic fern Ceratopteris pteridoides as revealed by AFLP analysis[J].Biochemical Systematics and Ecology,2010(38):1129-1136.
[73] Ren M X,Zhang Q G.Clonal diversity and structure of the invasive aquatic plant Eichhornia crassipes in China[J].Aquatic Botany,2007,87(3):240-246.
[74] Li X,Fan X,Chu H,et al.Genetic delimitation and population structure of three Trapa taxa from the Yangtze River,China[J].Aquatic Botany,2017,136(1):61-70.
[75] Liao Y Y,Yue X L,Guo Y H,et al.Genotypic diversity and genetic structure of populations of the distylous aquatic plant Nymphoides peltata (Menyanthaceae) in China[J].J.System.Evol,2013,51(5):536-544.
[76]Cao Q J,Mei F F,Wang L.Population genetic structure in six sympatric and widespread aquatic plants inhabiting diverse lake environments in China[J].Ecol Evol,2017(7):5713– 5723.
[77] 张光富,高邦权.江浙莼菜遗传多样性和遗传结构的ISSR分析[J].湖泊科学,2008,20(5):662-668.
[78] 刘朝贵,李小孟,须建,等.三大产区莼菜遗传多样性及亲缘关系的RAPD分析[J].西南大学学报(自然科学版),2012,34(10):54-59.
[79]Li Z Z,Gichira A W,Wang Q F,et al.Genetic diversity and population structure of the endangered basal angiosperm Brasenia schreberi (Cabombaceae) in China[J].Peer J,2018,6(5):e5296.
[80]Chen Y,Fan X,Li Z,et al.Low level of genetic variation and restricted gene flow in water lily Nymphaea tetragona populations from the Amur River[J].Aquatic Botany,2017(140):55-61.
[81] Yue X,Chen J,Guo Y,et al.Population genetic structure of Sagittaria natans (Alismataceae),an endangered species in China,revealed by nuclear SSR loci analyses[J].Biochemical Systematics and Ecology,2011,39(4-6):412-418.
[82] Wang Y,Chen J,Xu C,et al.Population genetic structure of an aquatic herb Batrachium bungei (Ranuculaceae) in the Hengduan Mountains of China[J].Aquatic Botany,2009,92(3):221-225.
[83]Zhang H Y,Tian K,Yu Y,et al.Genetic diversity among natural populations of Ottelia acuminata (Gaghep.) Dandy revealed by ISSR[J].Afr.J.Biotechnol.2009(8):6089–6093.
[84]Long C,Jiang Z,Dao Z.Genetic diversity of Ottelia acuminate (Hydrocharitaceae) from the Eastern Himalayas,revealed by ISSR markers[J].Bot.Orient.J.Plant Sci.,2010(7):56–63.
[85]Zhai S,Yin G,Yang X,et al.Population genetics of the endangered and wild edible plant Ottelia acuminata in Southwestern China using novel SSR markers[J].Biochem.Genet.2018(56):235–254.
[86] Li Y,Yu Y,Zhang X,et al.Genetic diversity of Ottelia acuminata var.crispa (Hydrocharitaceae):an endangered aquatic herb with extremely narrow distribution[J].Agric.Sci.J.Yanbian Univ.2016(38):139–148.
[87] Li Z,Lu M,Gichira A,et al.Genetic diversity and population structure of Ottelia acuminata var.jingxiensis,an endangered endemic aquatic plant from southwest China[J].Aquat.Bot.,2019(152):20–26.
[88]Fan X,Henry Kariuki Njerie,Wei Li,et al.Abundant historical gene flow within and among river systems for populations of?Ottelia acuminata var jingxiensis,an endangered macrophyte from southwest China[J].Aquatic Botany,2019 (157):1-9.
[89] Chen Y,Li X,Yin L,et al.Genetic diversity of the threatened aquatic plant Ottelia alismoides in the Yangtze River[J].Aquatic Botany,2008(88):10-16.
[90] 付春霖,戴小康,李小燕,等.黑藻cpDNA分子标记的筛选及其遗传多样性的初步研究[J].植物科学学报,2017,35(1):87-92.
[91] Wang B,Song Z,Liu G ,et al.Comparison of the extent of genetic variation of Vallisneria natans and its sympatric congener V.spinulosa in lakes of the middle–lower reaches of the Yangtze River[J].Aquatic Botany,2010,92(4):230-238.
[92]Wu Z,Yu D,Wang Z,et al.Great influence of geographic isolation on the genetic differentiation of Myriophyllum spicatum under a steep environmental gradient[J].Scientific Reports,2015(5):15618.
[93] Han Q,Wang G,Li W,et al.Genetic diversity of Potamogeton pectinatus L.in relation to species diversity in a pair of lakes of contrasting trophic levels[J].Biochemical Systematics and Ecology,2014(57):60-66.
[94] Li W,Xia L,Li J,et al.Genetic diversity of Potamogeton maakianus in the Yangtze River[J].Aquatic Botany,2004(80):227–240.
[95] 陈媛媛,栗琪,吴文颖,等.竹叶眼子菜居群遗传多样性和克隆结构[J].应用生态学报,2006(17):2034-2040.
[96]Chen Y,Li X,Yin L,et al.Genetic diversity and migration patterns of the aquatic macrophyte Potamogeton malaianus (Potamogetonaceae) in a potamo-lacustrine system[J].Freshwater Biology,2009(54):1178-1188.
[97] 陈锦华,汪小凡,梁述平.矮慈姑一个自然居群的遗传多样性[J].华中农业大学学报,2003,22(4):344-347.
[98]Poiani K A,Johnson W C.Evaluation of the emergence method in estimating seed bank composition of prairie wetlands[J].Aquatic Botany,1988(32):91-97.
[99] 李伟,刘贵华,周进,等.淡水湿地种子库研究综述[J].生态学报,2002,22(3):395-402.
[100] 刘贵华,萧蒇,陈漱飞,等.土壤种子库在长江中下游湿地恢复与生物多样性保护中的作用[J].自然科学进展,2007,17(6):741-747.
[101] 周进,Hisako T,李伟,等.受损湿地植被的恢复与重建研究进展[J].植物生态学报,2001,25( 5):561-572.
[102] 刘贵华.长江中下游湿地的种子库研究 [D].武汉:中国科学院研究生院(中国科学院武汉植物园),2005.
[103] Liu G H,Luo L M,Wang B,et al.Comparison of genetic variation in populations of wild rice,Oryza rufipogon,plants and their soil seed banks[J].Conserv.Gen.,2006,7(6):909-917.
[104]Pitman N C A,Jorgensen M J.Estimating the size of the world’s threatened flora [J].Science,2002(298):9891.
[105] 赵素婷.水鳖科两种水生植物种子活力维持机制研究[D].武汉:中国科学院大学(中国科学院武汉植物园),2018.
[106]Probert R J,Longley P L.Recalcitrant seed storage physiology in three aquatic grasses (Zizania palustris,Spartina anglica and Porteresia coarctata)[J].Ann.Bot.,1989 (63):53-63.
[107] Li E,Li W,Liu G,et al.The effect of different submerged macrophyte species and biomass on sediment resuspension in a shallow freshwater lake[J].Aquatic Botany,2008,88(2):121-126.
[108] Liu G,Zhou J,Li W,et al.The seed bank in a subtropical freshwater marsh:implications for wetland restoration[J].Aquatic Botany,2005,81(1):1-11.
[109] Zhang Y,Yin L,Jiang H,et al.Biochemical and biophysical CO2 concentrating mechanisms in two species of freshwater macrophyte within the genus Ottelia (Hydrocharitaceae)[J].Photosynthesis Research,2014(121):285-297.
[110]Shao H,Brigitte Gontero,Maberly S,et al.Responses of Ottelia alismoides,an aquatic plant with three CCMs,to variable CO2 and light[J].Journal of Experimental Botany,2017(68):3985–3995.
[111] 于丹,种云霄,涂芒辉,等.中国水生高等植物受危种的研究[J].生物多样性,1998,6(1):13-21.
[112] 赵魁义,何舜平,李伟,中国湿地生物多样性研究[J].中国科学院院刊,2010,25(6):659-667.





