摘 要: 以科尔沁沙地为研究对象, 采用方差分析和回归分析的方法研究沙地群落的物种组成及生物多样性.结果表明:1) 从流动到固定沙丘, 群落的物种数从4. 533增加到9. 033, 增加了99. 27%.群落盖度、高度、密度和地上生物量分别增加了370. 1%、171. 6%、156. 1%和526. 7%, 其中流动沙丘与半流动沙丘 (或半固定沙丘) 、固定沙丘之间的盖度、高度、密度差异显着 (P <0. 05) . 2) 科尔沁沙地植被中均以一年生植物占比最高, 但沙地从流动沙丘演变到固定沙丘, 一年生植物的优势度下降了25. 48%, 多年生植物的优势度增加了92. 37%. 3) 物种多样性与均匀度指数随着沙地的恢复波动变化, 在半固定沙丘最大;而群落的物种丰富度呈上升趋势, 固定沙丘最大.
关键词: 沙地群落; 物种组成; 生物多样性;
Abstract: Taking Horqin Sandy Land as the research object, the species composition and biodiversity of sandy communities were studied by means of analysis of variance and regression analysis. The main conclusions were as follows: 1) From the shifting sandy dune to the fixed sandy dune, the number of species in the community increased from 4. 533 to 9. 033, an increase of 99. 27%. Community coverage, height, density, and above-ground biomass increased by 370. 1%, 171. 6%, 156. 1% and 526. 7%, respectively, and there was a significant difference between shifting sandy dune and semi-shifting sandy dune ( or semi-fixed sandy dune) and fixed sandy dune ( P < 0. 05) . 2) The Horqin sandy land communities were dominated by annual plants. However, with the restoration of communities, the dominance of annual plants showed a downward trend, which dropped by 25. 4%, and perennial plants showed an upward trend, which increased by109. 39%. 3) From shifting sandy dune to fixed sandy dune, the species diversity and evenness of the communities first increased and then decreased, and reached the maximum in semi-fixed sandy dune, and the species richness of the communities gradually increased.
Keyword: sandy communities; species composition; biodiversity;
科尔沁沙地处于东北平原和内蒙古高原之间的过渡区域, 同时也是农牧交错区的脆弱地带, 是典型的半干旱区[1].由于过度开垦、超载放牧以及其他各种因素的影响, 导致植被减少、水土流失严重、生态环境恶化.近年来, 通过采取了禁牧等措施, 沙地恢复成效显着.群落的物种组成及生物多样性反映了群落和生态系统的结构类型、组织水平、发展阶段、稳定程度和生境差异等, 因此研究植物群落的特征和多样性于退化生态系统的恢复有着重要的指导意义[2,3].金艳霞等研究了不同年代取土场高寒草原植物群落在自然恢复过程中的生产力、物种多样性、综合恢复能力和稳定性动态以及土壤养分变化等[4].颜衡祁探讨了衡阳紫色土丘陵坡地恢复过程中植物群落结构及多样性的变化[5].肖玉等对青藏苔草高寒草原退化序列进行研究分析, 解析了高寒草原退化过程中植物群落数量及表观特征的变化过程[6].Danie和John研究爱荷华州中西部的6个恢复地点和3个附近的剩余高草草场, 发现恢复种植包含早期物候物种的种子混合物能提高生物多样性[7].吴旭东等人探讨了沙漠化草地及灌丛化草地土壤有机碳稳定性与土壤性状及草地群落间的相关性[8].但是关于科尔沁沙地四种沙丘类型的植物群落组成及数量特征变化的研究有限, 故本文中研究不同恢复阶段中科尔沁沙地群落的物种组成、群落特征及生物多样性的动态变化, 旨在为退化沙地植被恢复与重建提供理论依据.

1、 研究区概况
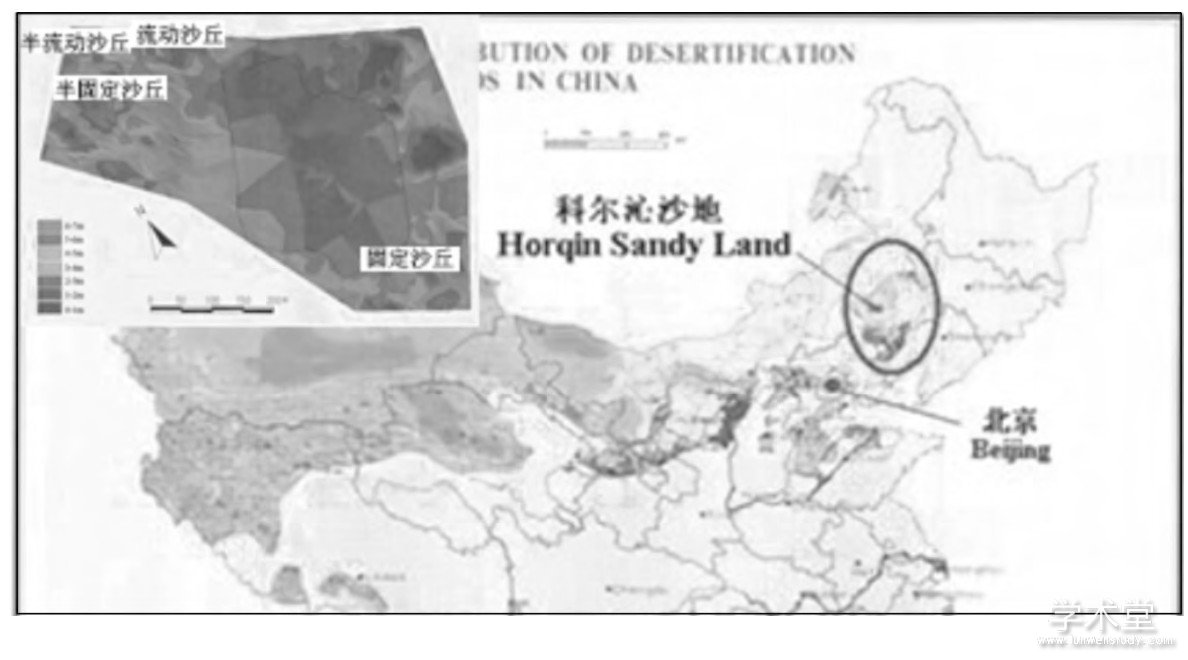
研究区位于科尔沁沙地中南部的奈曼旗中国科学院奈曼沙漠化研究站境内 (42°54'N, 120°41'E) (图1) .该区属半干旱气候, 年平均降水量366 mm, 年蒸发量1 935mm, 年均气温6.5℃, 1月平均气温-12.7℃, 7月平均气温23.7℃, ≥10℃积温3 000℃以上, 无霜期150 d[9].土壤类型为沙质栗钙土, 经破坏后则退化为流动风沙土, 沙土基质分布广泛, 风沙活动强烈[10].
图1 科尔沁沙地地理位置示意图
4种类型的沙地群落的特征各不同, 流动沙丘群落:沙丘表面几乎裸露, 无结皮或仅有少量物理性结皮, 植被稀松, 盖度小于20%, 主要植被有差巴嘎蒿 (Artemisia halodendron) 、沙地沙米 (Agriophyllum squarrosum) 和狗尾草 (Setaria viridis) 等;半流动沙丘群落:流沙面积大于10%, 地表已有结皮, 但数量较少, 群落盖度在20%~60%之间, 主要植被是狗尾草 (Setaria viridis) 、大果虫实 (Corispermum macrocarpum) 和差巴嘎蒿等;半固定沙丘群落:地表及植被盖度与半流动沙丘相似, 优势物种发生变化, 以三芒草 (Aristida adscensionis) 、猪毛菜 (Salsola collina) 和差巴嘎蒿为主;固定沙丘群落:流沙面积在10%以下, 地面覆盖固定风沙土或沙质栗钙土或已形成较厚的土壤结皮, 群落盖度大于50%, 主要植被有达乌里胡枝子 (Lespedeza davurica) 、虎尾草 (Chloris virgata) 以及糙隐子草 (Cleistogenes squarrosa) 等.
2、 研究方法
2.1、 野外调查方法
根据空间替代时间的原则, 2014年8月, 在野外选分别处于流动沙丘、半流动沙丘、半固定沙丘和固定沙丘等不同恢复阶段的4个演替时间不等的典型群落样地进行调查 (图2) , 在每个样地的典型地段等距离 (20 m, 南北向) 设置3条长100 m的样带, 在每条样带上等距离 (10 m) 设置10个固定样点, 一个样地30个样点, 共计120个样点, 在每个取样点的旁边, 设置面积1 m2的草本样方, 测定每个样方的种类组成及其特征值 (盖度、高度和密度) , 采用齐地面刈割法调查地上生物量, 草样在85℃烘干24 h称重.各样地每种植物的出现频率根据30个样点的测定数据计算.
2.2、 数据处理
2.2.1、 物种重要值和优势度
物种的重要值和优势度根据群落的相对密度、相对频度、相对高度和相对盖度进行计算.
图2 采样点示意图
IV=相对密度+相对频度+相对高度+相对盖度, DS=IV/4×100.
式中, IV是重要值 (Important value) , DS为优势度 (Dominance of species) .
2.2.2、 物种多样性
采用Patrick指数、Simpson指数、Shannon-wiener指数和Pielow均匀度指数进行衡量.
式中S为物种总数, Pi为第i种植物的个体数占群落中总个体数的比例.
2.3、 统计分析
本文中采用方差分析比较各个群落特征之间的差异, 数据统计分析均在Excel和SAS9.4中完成.
3、 结果与分析
3.1、 沙地群落的基本特征
表1是4种植物群落的物种数、盖度、高度、密度和地上生物量.从流动到固定沙丘, 群落的平均物种数由4.533到9.033, 增加了99.27%.群落总盖度、平均高度、平均密度和地上生物量的增幅分别是370.1%、171.6%、156.1%和526.7%.其中流动沙丘与半流动沙丘 (或半固定沙丘) 、固定沙丘之间的盖度、高度和地上生物量差异显着 (P<0.05) , 但是半流动与半固定沙丘之间差异不明显;流动沙丘 (或半流动沙丘) 与固定沙丘之间的密度差异显着 (P<0.05) , 但流动沙丘、半流动和半固定沙丘之间差异不显着.
表1 沙地植物群落的数量特征 (均值±标准误)
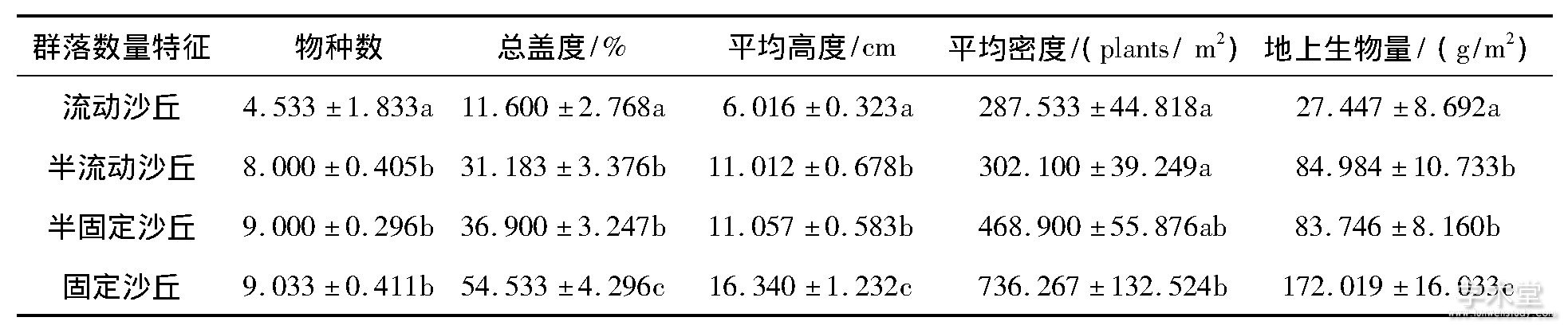
同一列数据内不同字母表示差异显着 (P<0.05)
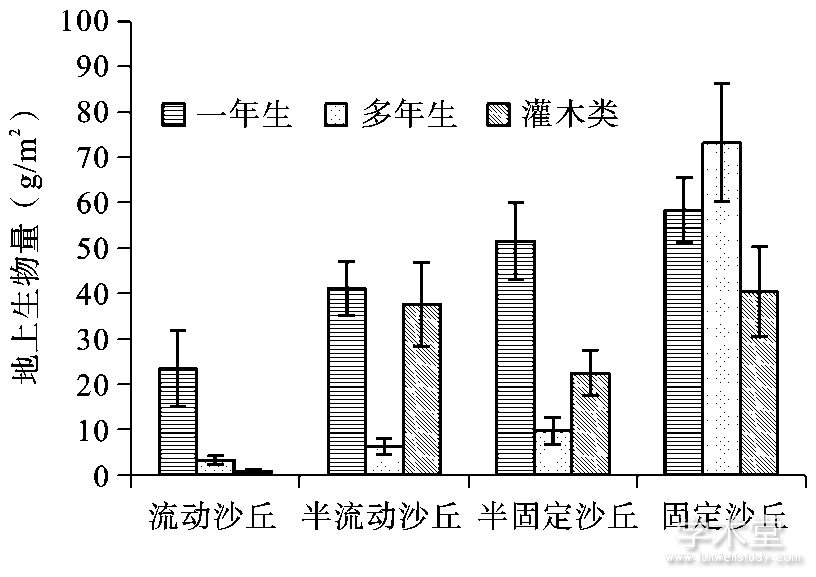
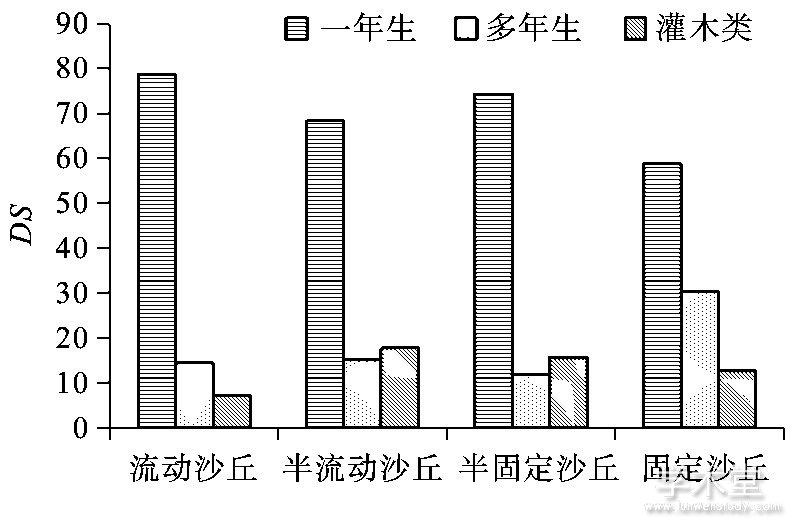
从流动沙丘到固定沙丘, 一年生植物地上生物量逐渐增加, 其占比在流动沙丘、半流动沙丘以及半固定沙丘均最大, 分别为85.47%、48.39%、61.58%.多年生植物地上生物量也呈上升趋势, 在固定沙丘中其占比最高, 为42.6%.灌木类地上生物量呈波动变化 (图3) .
图3 4种沙地类型不同生活型地上生物量
3.2 沙地群落的物种优势度
物种优势度是用来表示某个种在群落中的地位和作用的综合数量指标, 优势度的变化体现了该物种在群落中作用的变化[11].沙地植被类型改变中, 群落的优势物种逐渐发生了变化, 流动沙丘的优势物种是大果虫实、狗尾草, 占比59.03%.半流动沙丘的优势物种是大果虫实、狗尾草和差巴嘎蒿, 占比58.20%.半固定沙丘的优势物种是狗尾草、三芒草、大果虫实等, 占比为43.86%, 并且该阶段新进驻了虎尾草、山竹岩黄芪 (Hedysarum fruticosum) 等物种, 而沙米等沙生植物退出群落.固定沙丘的优势物种是画眉草 (Eragrostis pilosa) 、黄蒿 (Artemisia scoparia) 、猪毛菜和达乌里胡枝子, 占比为44.70% (表2) .
流动沙地到固定沙地的变化中, 从科属组成来看, 4个群落均以禾本科、藜科、豆科和菊科为主.禾本科和豆科植物优势度逐渐增加, 分别增加了18.84%和429.03%, 藜科植物优势度逐渐下降, 降低了93.87%, 而菊科植物的优势度先增加后降低, 主要是受优势物种差巴嘎蒿的影响.
表2 沙地群落物种优势度
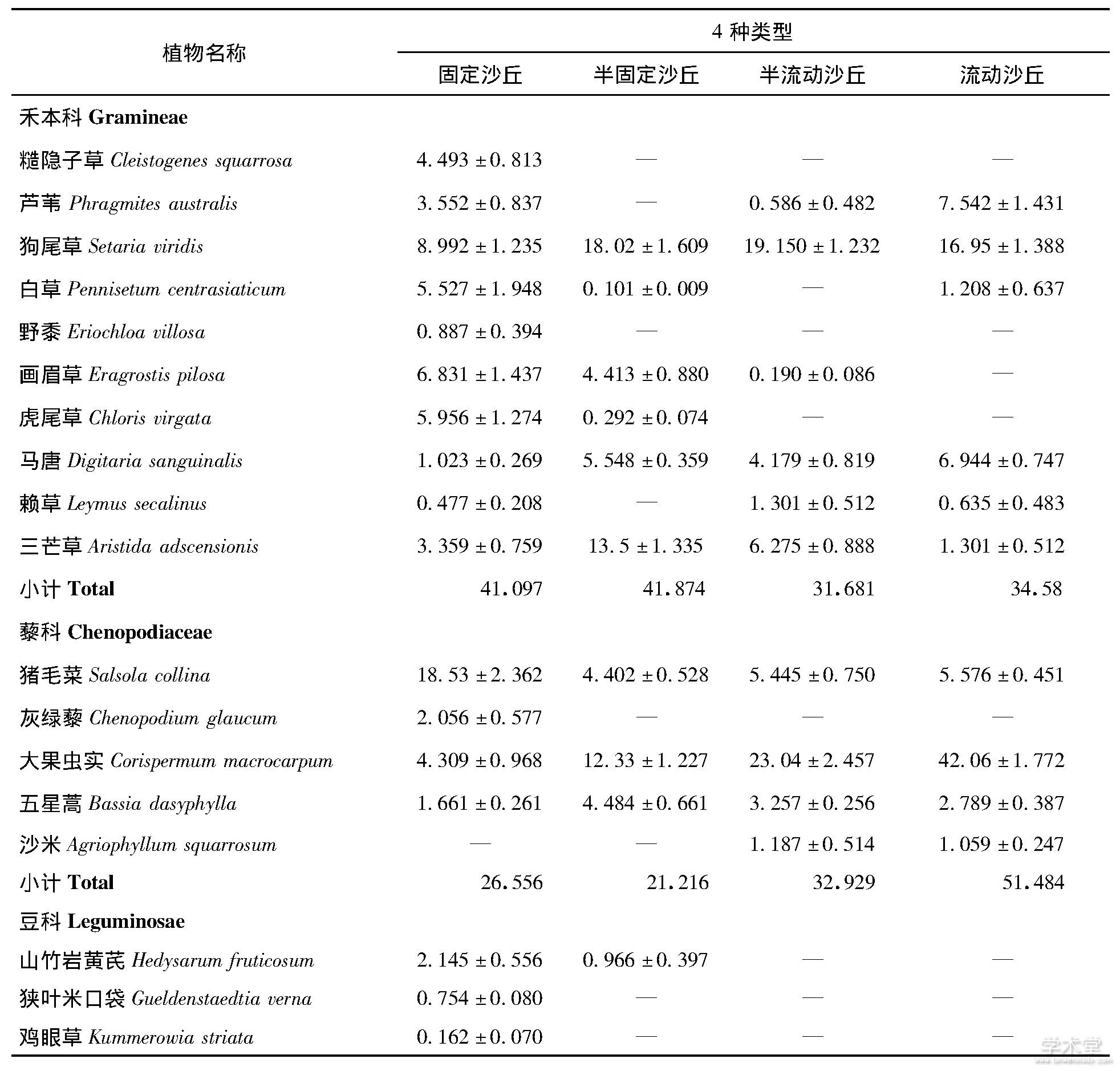
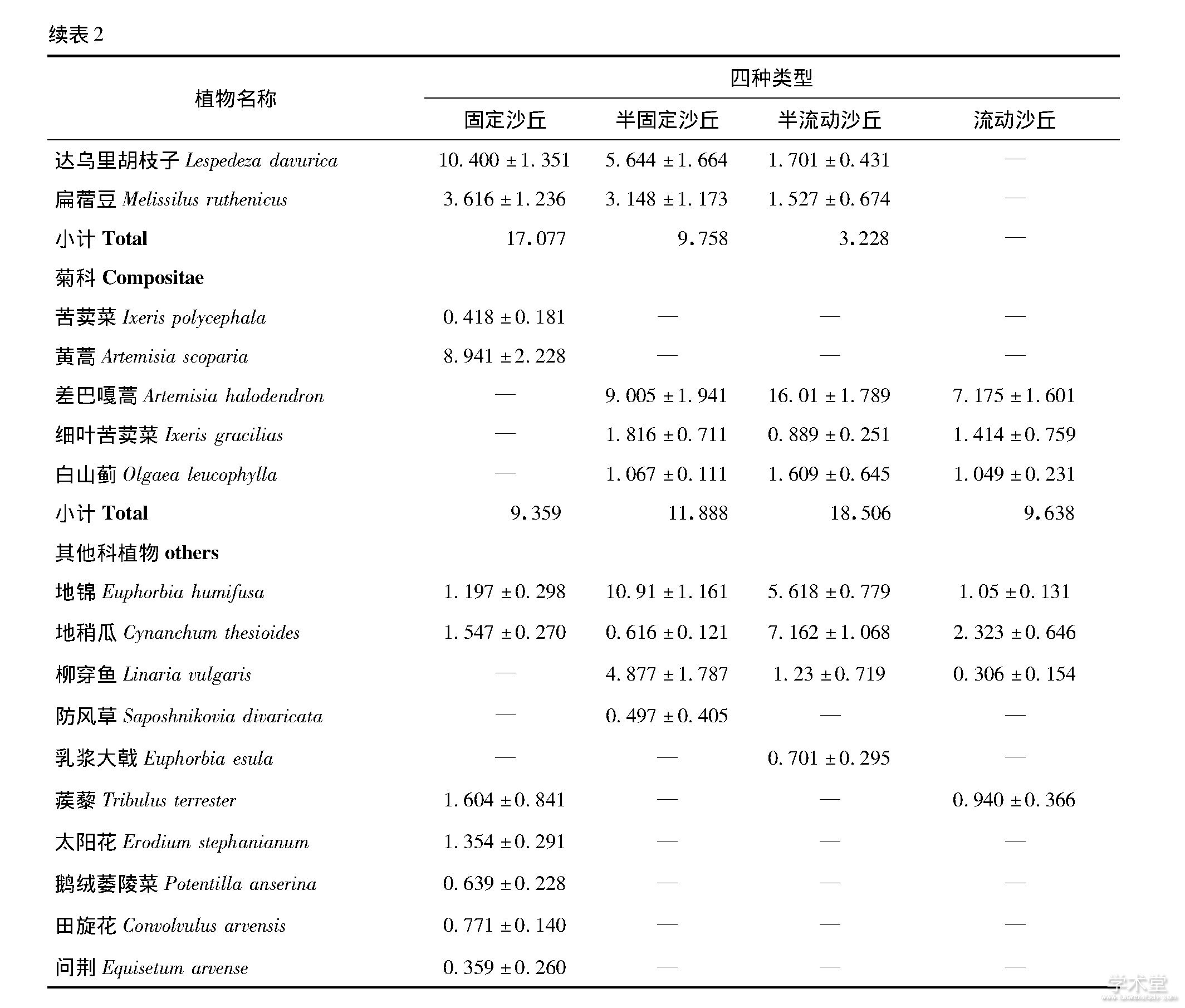
图4 不同生活型物种优势度
从生活型来看 (图4) , 群落优势度均以一年生植物为主, 但从流动沙丘到固定沙丘, 一年生植物的优势度逐渐下降, 下降比例为25.48%.大果虫实、五星蒿 (Bassia dasyphylla) 和狗尾草是沙地群落的常见物种, 出现在各个群落中;沙米作为沙地的先锋植物, 流动沙丘优势度最大, 到半固定沙丘退出群落;画眉草、虎尾草、野黍 (Eriochloa villosa) 及鸡眼草 (Kummerowia striata) 等植物逐渐进驻群落, 多年生植物糙隐子草、白草 (Pennisetum centrasiaticum) 和狭叶米口袋 (Gueldenstaedtia verna) 、扁蓿豆 (Melissilus ruthenicus) 等的优势度逐渐增加, 从流动到固定沙丘, 多年生植物的优势度增加了109.39%.灌木类的优势度先增加后降低, 差巴嘎蒿的优势度在半流动沙丘最大, 半固定沙丘次之, 直至固定沙丘退出群落.
3.3、 沙地恢复群落的物种多样性
沙地群落的Simpson指数和Shannon-Weiner指数从流动沙丘到固定沙丘逐渐增加, 在半固定沙丘达到最大, 之后降低, 且流动沙丘与半流动 (或半固定沙丘) 、固定沙丘之间差异显着 (表3, P<0.05) .群落的Patrick指数 (物种丰富度) 逐渐增加, 从半流动到固定沙丘, 物种丰富度差异不显着 (P<0.05) .群落的Pielow均匀度指数先增加后降低, 固定沙丘最低.群落物种多样性波动变化, 这不仅与物种组成有关, 还与群落内部物种组成的均匀度等有关系.
表3 沙地群落物种多样性
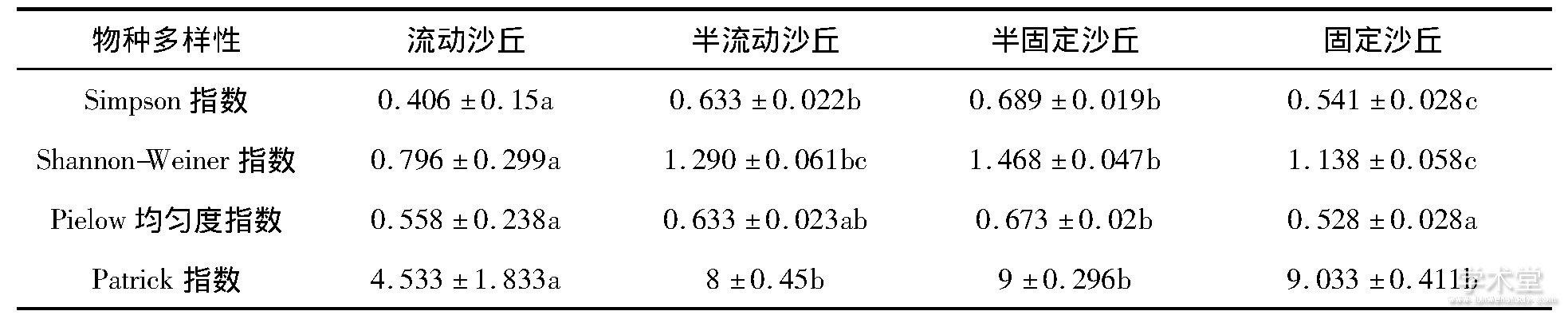
同一列数据内不同字母表示差异显着 (P<0.05)
4、 讨论
流动沙丘经封育后逐渐演变成半流动、半固定和固定沙丘, 随之植物群落的密度、高度、盖度和地上生物量均显着提高.这与罗永清等人的研究结果一致, 流动沙丘的密度、盖度、生物量均显着低于固定沙丘[12].王国梁等人的研究中指出, 短期围封能有效提高群落的数量特征, 中后期围封在提高群落数量特征的同时还使群落优势物种发生更迭[1,2,3].科尔沁沙地植被主要由禾本科、菊科、豆科和藜科组成, 随着群落的恢复, 禾本科与豆科植物在群落中地位上升, 藜科植物的作用逐渐降低.这与左小安等人的研究结果相似, 即科尔沁沙地藜科植物优势度则随着沙丘固定和植被恢复波动下降, 豆科植物优势度波动变化, 菊科植物主要是差巴嘎蒿, 禾本科植物在沙丘固定和植被恢复过程中物种数和优势度增加[14].
沙地植被演替进程中, 物种组成与群落演替的动态变化格局反映了生态系统恢复过程中群落环境的变化和生物多样性对这种变化的响应过程[15].流动沙丘由于植被盖度低, 导致地面水分蒸发量较大, 水土流失严重, 土壤养分低, 反而不利于植物的生长[16].因此该阶段群落的密度、地上生物量等均较低, 由于缺乏植被的保护, 风沙活动强烈, 基质条件极不稳定, 不利于植物繁殖体的传播和定居[17].只有适应性强、繁殖速度快的植物能够生存下来, 如沙米、大果虫实、狗尾草等, 一年生植物在流动沙丘占据绝对优势.正如张志南的研究认为, 处于半干旱区的植被结构及优势物种极大程度受土壤水分的影响, 例如杂草类植物在生境条件艰巨的区域占比更高[18].在半流动沙丘, 随着群落的演替和植物的生长, 沙地植被的盖度明显提升, 并且生态系统功能加强, 使得多年生植物及灌木类获取更多的生长机会, 也增加了群落的地上生物量, 同时群落中物种对限制性资源的竞争增强[8].因此竞争力较强的植物如差巴嘎蒿、大果虫实、狗尾草成为优势物种.差巴嘎蒿极大地提高了半流动沙地群落的优势度和地上生物量, 其枝叶也增加了植被的盖度.群落环境得到改善, 对土壤表面的保护作用加强, 降低风蚀, 拦截降尘, 使细粒物质沉积, 土壤结皮逐渐形成, 沙丘演变成半固定状态.在半固定沙丘, 狗尾草、三芒草、大果虫实成为优势物种, 由于植被发育增加了土壤基质的稳定性, 更多植物物种的繁殖体能够扩散和定居.同时细颗粒物质沉积以及有机质积累改善了土壤养分条件:细沙粒及粉、黏粒含量、表层土壤中的速效P含量、全C、全N含量均增加[19].此时更有利于一年生禾本科植物的生长, 群落的物种组成及结构得到优化.有研究表明随着植被的恢复, 维持植被发育的环境条件进一步改善, 许多物种的繁殖体实现成功的散布和定居[20].因此在固定沙丘, 生态条件达到“最优”, 群落的地上生物量大幅度提升, 盖度、密度等也显着增加, 土壤肥力也不断增加, 多年生草本植物逐渐取代了一年生草本植物, 画眉草、虎尾草、猪毛菜和达乌里胡枝子等成为科尔沁固定沙丘群落的优势物种.
群落物种多样性与很多因素有关, 一般认为随着群落的恢复演替, 物种多样性会逐渐增加.但是关于物种多样性指数与恢复演替关系的研究说法不一, 有的学者认为物种丰富度指数随恢复演替过程呈现出抛物线型, 也有的学者发现多样性指数随演替进程先高后低, 呈波浪式降低[21].对于科尔沁沙地, 物种多样性与均匀度随着群落的演替先增加后降低, 与大多学者所提出的“中等干扰假说”一致[22].张华的研究表明从半流动沙地恢复到半固定沙地阶段, 物种多样性恢复最快, 且随土壤粗颗粒含量的减少、细微颗粒含量和土壤养分含量的增加, 植物多样性指数提高[19].
5、 结论
1) 从流动到固定沙丘, 群落的物种数从4.533增加到9.033, 增加了99.27%.群落盖度、高度、密度和地上生物量分别增加了370.1%、171.6%、156.1%和526.7%, 流动、半流动 (半固定) 、固定沙丘三者之间的数量特征差异明显 (P<0.05) .
2) 从流动到固定沙丘, 一年生植物的优势度下降了25.4%, 多年生植物增加了109.39%.其中, 流动沙丘的优势物种是大果虫实、狗尾草;半流动沙丘的优势物种是大果虫实、狗尾草和差巴嘎蒿;半固定沙丘的优势物种是狗尾草、三芒草、大果虫实等;固定沙丘的优势物种是画眉草、虎尾草、猪毛菜和达乌里胡枝子.禾本科、豆科植物优势度分别增加了18.84%和429.03%, 藜科植物降低了93.87%.
3) 群落的物种多样性和均匀度指随着群落恢复逐渐增加, 在半固定沙丘达到最大, 之后降低, 其中Simpson指数在0.4060.689之间, Shannon-Weiner指数在0.7961.468之间, 均匀度指数在0.5280.673之间.群落的丰富度呈上升趋势, 固定沙丘达到最大, 丰富度为9.033.
参考文献
[1]张婧, 左小安, 吕朋, 等.科尔沁沙地草地植被对围封和放牧的响应[J].中国沙漠, 2017 (3) :446-452.
[2]史作民, 程瑞梅, 刘世荣, 等.宝天曼植物群落物种多样性研究[J].林业科学, 2002, 38 (6) :17-23.
[3]阎海平, 谭笑, 孙向阳, 等.北京西山人工林群落物种多样性的研究[J].北京林业大学学报, 2001, 23 (2) :16-19.
[4]金艳霞, 周华坤, 姚步青, 等.高寒草原取土场自然恢复过程中植物群落的特征和恢复力[J].草业科学, 2014, 31 (8) :1528-1537.
[5]颜衡祁.衡阳紫色土丘陵坡地恢复过程中植物群落结构及多样性的变化[J].中国农学通报, 2015, 31 (19) :180-184.
[6]肖玉.青藏高原高寒草原不同退化程度植物群落特征与土壤养分的关系[D].兰州:兰州大学, 2016.
[7]Carter D L, Blair J M. Recovery of native plant community characteristics on a chronosequence of restored prairies seeded into pastures in West-Central Iowa[J]. Restoration Ecology, 2012, 20 (2) :170-179.
[8]吴旭东.沙漠化对草地植物群落演替及土壤有机碳稳定性的影响[D].银川:宁夏大学, 2016.
[9]赵丽娅, 高丹丹, 熊炳桥, 等.科尔沁沙地恢复演替进程中群落物种多样性与地上生物量的关系[J].生态学报, 2017, 37 (12) :4108-4117.
[10]刘新民, 赵哈林, 赵爱芬.科尔沁沙地风沙环境与植被[M].北京:科学出版社, 1996.
[11]郑秋红, 伍永秋, 张宏, 等.怀来盆地丘陵区弃耕地自然恢复中植物群落特征的变化[J].北京师范大学学报 (自然科学版) , 2005 (6) :627-631.
[12]罗永清, 赵学勇, 丁杰萍, 等.科尔沁沙地不同类型沙地植被恢复过程中地上生物量与凋落物量变化[J].中国沙漠, 2016, 36 (1) :78-84.
[13]王国梁, 刘国彬, 刘芳, 等.黄土沟壑区植被恢复过程中植物群落组成及结构变化[J].生态学报, 2003 (12) :2550-2557.
[14]左小安, 赵哈林, 赵学勇, 等.科尔沁沙地不同恢复年限退化植被的物种多样性[J].草业学报, 2009, 18 (4) :9-16.
[15]曹成有, 蒋德明, 朱丽辉, 等.科尔沁沙地草甸草场退化的原因与植物多样性变化[J].草业学报, 2006, 15 (3) :18-26.
[16]杨宁, 邹冬生, 李建国.衡阳盆地紫色土丘陵坡地植被恢复模式建设[J].草业科学, 2010, 27 (10) :10-16.
[17]刘志民, 蒋德明, 阎巧玲, 等.科尔沁草原主要草地植物传播生物学简析[J].草业学报, 2005, 14 (6) :23-33.
[18]张志南, 武高林, 王冬, 等.黄土高原半干旱区天然草地群落结构与土壤水分关系[J].草业学报, 2014, 23 (6) :313-319.
[19]张华, 伏乾科, 李锋瑞, 等.退化沙质草地自然恢复过程中土壤一植物系统的变化特征[J].水土保持通报, 2003 (6) :1-6.
[20] 曹成有, 蒋德明, 朱丽辉, 等.科尔沁沙地草甸草场退化的原因与植物多样性变化[J].草业学报, 2006, 15 (3) :18-26.
[21]韦兰英, 韦启忠, 莫祝平, 等.不同干扰方式对桂西北灌草植被物种多样性和生产力的影响[J].水土保持研究, 2016, 23 (4) :288-293, 299.
[22]杨宁, 邹冬生, 李建国, 等.衡阳盆地紫色土丘陵坡地主要植物群落自然恢复演替进程中种群生态位动态[J].水土保持通报, 2010, 30 (4) :87-93.





