����ժ Ҫ�������о�ּ��̽������ģ������ЧӦ�о������Թ�Ӭ�˶���˯��Ӱ��Ŀ�����.ͨ�������ܹ���ģ������������ʵʱ����Ӭ��Ϊ�������λ�ǣ�����ʱ�䣨3 d��ģ���������������У�����ʱ�䣨10 d��20 d��30 d����������Ӭ�˶���˯�ߵı仯��ѡȡ��Ӱ������ŵĶ�ʱ�䴦���飬�о�ģ������ЧӦ�������Ӻ��Ļ���period (per����timeless(tim����clock (clk����cycle (cyc����cryptochrome (cry���������ʶ�Ͱ���dopamine,DA����5-��ɫ����5-hydroxytryptamine,5-HT���ؼ��ϳ�ø���������ø���Ұ����ǻ�ø��ɫ�����ǻ�ø���ı������ddc��pale��trh����ˮƽ��DA��5-HT������Ӱ��.�����ʾ����ʱ�䱩¶�£���Ӭҹ�����˶������ӡ���λʱ���˶��������ӡ�˯��ʱ��ʹ������١������ӻ���tim��clk��cyc��cry�����ʺϳ���ر������ddc��pale��trh�ı���ˮƽ��������������ʱ�䴦�������Ӭ�˶���˯�ߵ�Ӱ���С.���о���Ϊ����ģ������ЧӦ�о������Թ�Ӭ�˶���˯�ߵ�Ӱ���ǿ��еģ�����о�����Ժ���ҽѧ�о����н������.
�����ؼ��ʣ���ģ������; �����λ��; ��Ӭ; �˶�; ˯��;
����Abstract����This study was aimed to investigate the feasibility of using simulated microgravity to study the effects of microgravity on activity and sleep of Drosophila melanogaster. Simulated microgravity experiments performed using the random positioning machine(RPM) and male Drosophila melanogaster, and the activity and sleep of the flies were monitored using the DAM system(Trikinetics, USA). First, flies were exposed to simulated microgravity for a short time(3 days) with DAM system monitoring. Next, after being exposed to simulated microgravity for a long time(10, 20, and 30 days), flies were removed to be monitored for 3 consecutive days under control conditions. Then, the flies after short-term treatment(3 days) with greater impact were sampled to study the effects of simulated microgravity on the major circadian clock genes period(per), timeless(tim), clock(clk), cycle(cyc), cryptochrome(cry), and ddc, pale, trh genes encoding dopa decarboxylase, tyrosine hydroxylase, tryptophan hydroxylase, that involved in synthesis of neurotransmitter dopamine(DA) and 5-hydroxy-tryptamine(5-HT), and the contents of DA and 5-HT. The results showed that during short-term treatment, total activity accounts and activity accounts in wake period both increased, and total sleep and number of sleep episode both decreased at night; after short-term treatment, the relative expression levels of tim, clk, cyc and cry increased, and the relative expression levels of ddc, pale and trh increased; after long-term treatment,overall effect on activity and sleep was slight. This study suggests that it is feasible to study the effects of microgravity on activity and sleep of Drosophila melanogaster by simulated microgravity, and valuable reference can be provided for aerospace medical research.
����Keyword����imulated microgravity; random positioning machine; Drosophila melanogaster; activity; sleep;
���������й����˺�����ҵ�IJ��Ϸ�չ��δ���й��ռ�վ�Ľ��ɲ�Ͷ��ʹ�ã����и���ĺ���Ա���칤��ʦ������̫�գ��������ڹ칤��.����о�����������Ա�ڹ��ڼ��ձ����˯��ʱ��̡�˯���������˯������.���õ�˯�߶����彡������Ҫ�Բ��Զ���[1,2,3]����α��Ϻ���Ա���ڹ������˯�߳�Ϊؽ���������Ҫ��ѧ����.�ռ价�����������⡢��������ҹ���ɼ���Ϣ�ƶ�[4,5]�ȱ���Ϊ����ɺ���Ա˯���ϰ�����Ҫ����Ϊ�˿�ѧ���ǴӺ���������ơ����ü�˯���������õ�����Ա��ѵ������չ�˴����Ĺ���.��ʵ�ʵ�������ʾ����Щ��������ȫ�������Ա���ڹ�˯������[4,5,6,7,8].��������Ŀǰ�Կռ价�����ر���������˯�ߵ�Ӱ�켰���û��Ƶ��˽���٣�������ԵضԺ���Ա����ѵ����������ʵ��������ɸ�Ԥ���������.
����Ŀǰ��������˯�ߵ�Ӱ���о���Ҫ�������ʵ��͵ػ�ģ��ʵ�飬ǰ����Ҫ���Թ��⺽��Ա�ķ��б��漰����ڹ������������ڵ��о������ʼ�����.���������������Һ����뺽�վֵĶ����κ���Ա�����ұ������¼�Ǽ������ʾ�����ຽ��Ա��̫���У�˯��ʱ�����Լ��٣���������6 h����ÿ��ҹ���IJ����ϴ�ͬʱ˯�߽ṹҲ�����˸ı䣬˯�����������Һ���Ա���ձ���ڴ�˯����ҩ�ļ�¼.��Ȼ���ú�������˯ʱ��������˯��������ߣ����嶯��¼����ʾ˯��Ч�ʲ�û����ߣ�˯��ʱ��Ҳ��û�����ӣ����Ҵ���ҩ��ĸ�����Ҳ�dz�ֵ�þ���[4,9,10].
�������ں����������������Ժͺ�����ؿռ���Դ��ϡȱ�ԣ����ú���Ա��ʵ�鶯�↑չ�ڹ��о������ڽϴ�����ѣ����Խ��н�Ϊϵͳ��������û����о�.ģ������ЧӦ�ĵػ�ʵ���Ϊ��Ҫ�ĸ����о��ֶΣ���ͷ��λ�Դ���[11,12]��β����[13]���֫ж��[14]���������λ��[15,16,17,18,19,20,21,22,23,24,25,26,27,28]��.���У������λ��ƾ�������ڲ����������������������ȶ��ɿ������ڶ����ڽ��ж���ظ������ƣ�����Խ��Խ�㷺.��Ŀǰ�ػ�ģ��ʵ���о���ȱ������ģ������ЧӦ��˯��Ӱ�����ر���������Ҫԭ��֮һ��ȱ����Ӧ���о��豸���о�ģ�ͺ����۷���.
������Ӭƾ�����С����ֳ�켰�����ڶ̵��ص�����Ƴ�Ϊ�˺�����ص�“����”�������о�������Ӭ�Կռ价����Ϊ���У�������л��������һ�����������Ŵ���Ӧ[27,28,29,30,31,32]����Ȼ�ػ���ʵ������������Ӭ�ܹ��������λ��ģ�������ЧӦ������Ӧ[19,20]���������λ�DZ�¶�Ĺ�Ӭ�ܹ�������������ʵ���������ƵĻ������[27,28].�����������λ���о���Ӭ��Ϊ���ر���˯����Ϊ�仯���о�����δ������.����������Ӭ��Ϊ�о�˯����Ϊ��ģʽ���ﱻ�㷺Ӧ��[33,34,35,36,37,38]�����о�����һ�������ڹ�Ӭ˯���о��������λ�ǣ����ӹ�Ӭ����ɺ������ӵ��ػ��Ƶı仯�����������һ����̽�����������λ���о���Ӭ˯����Ϊ�Ŀ����ԣ���һ���棬����̽��ģ������ЧӦ�Թ�Ӭ˯�ߵ�Ӱ�켰���û��ƣ��Ӷ�Ϊ�о�����ЧӦ��������˯�ߵ�Ӱ���ṩ�µ�˼·.
����1�� �����뷽��
����1.1 ����Ӭ����
����Ұ���ͺڸ���Ӭ��Drosophila melanogaster)Canton-S(CS��Ʒϵ�������й���ѧԺ�Ϻ�������ѧ�о�Ժ����ϸ������Ӭ��Դ�뼼��ƽ̨.��Ӭ������������-��-��ĸ���������˹��������У���������12 L��12 D(6��00 am���ƣ�6��00 pm�صƣ����¶ȣ�25±1���棬���ʪ�ȣ�60±2��%.���о����ù�Ӭ��ΪCS���Թ�Ӭ.
����1.2 ������ЧӦģ��ϵͳ
�������о����õ�����ЧӦģ��ϵͳ���й���ѧԺ���л����о������Ϻ����������о�����Թ�Ӭ���ص��������ƣ���Ҫ�������λ�ǣ�ͼ1a����ϵͳ���ƣ�ͼ1b��c�����������.�����λ����Ҫ�ɻ�������ֱ֧�żܡ���ת����������ת������������ϵͳ����Ʒ��װ��Ԫ��Դ���.������Ʒ��װ��Ԫ��Թ�Ӭ��Ϊ����豸DAM2�������DPM�������TriKinetics�����������ص㣬�������Ӧ����Ʒ̨����ص�Դ���źŴ�����ö��Ƶ��紫�������ƣ��ܹ����������ת���������Ť�ˣ���֤�豸�ȶ�����ͬʱ��ʵ�ּ�����ݵ�ʵʱ����.��Դ��LED������ɣ����ԶԹ���ʱ�估ǿ�Ƚ����ȶ���ȷ�Ŀ���.ϵͳ�����ɵ���䣨ͼ1b���Ϳ���������ͼ1c����ɣ���������ģ�ⵥԪ�ŷ����������ϵͳ��ͬʱ���ݹ�Ӭ���ص㼰������ױ�����������ڹ�Ӭ�о��Ļ�ת������ģ������ʾ����ģ��Ч����ԼΪ10-4g.
����Fig.1 Microgravity simulation system

����(a)Random positioning machine.(b)Electric cabinet.(c)Control software.
����1.3�� �˶���˯����Ϊ���
����1.3.1 ����ʱ��ģ������ЧӦ
������������Ӭװ���Ӭ��Ϊ���ϵͳDAM2�����������TriKinetics���У�����������£��ڼ�ⲣ���ܣ��ھ�Ϊ3.5 mm������Ϊ65 mm��һ��װ����������5%����+2%��֬������Ӭʳ�ã���һ����������ס�ܿڣ�ÿֻ������װ��һֻ��Ӭ��ÿ���������32�����ܣ�ÿ5 min��¼һ����Ӭ���˶����.���ô����飨G�飩�Լ������飨C�飩����G��ļ���������������λ���У���������24 h����ʹ��Ӭ��Ӧ���ڵ�3��6��00 am�����豸��������3 d��֮��ر��豸������������������24 h���Թ۲���Ӭ�Ļָ������C��ļ���������������λ���ԣ�ȷ�����������Ĺ��ա��¶ȡ�ʪ�ȵȻ�����ͬ.
����1.3.2 ����ʱ��ģ������ЧӦ
�����ڶ�ʱ��ģ������ЧӦ����Ӭ�˶���˯�ߵ�Ӱ���о��У��豸�رպ���Ӭ���˶���˯����Ϊ�ܿ�ָ�������״̬�����2.1.1��.��ˣ���չ�˽ϳ�ʱ��Ĵ������������£���������Ӭװ���Թ��У����ô����飨G�飩�Լ������飨C�飩����G����Թܷ����������λ���У���������24 h����ʹ��Ӭ��Ӧ�价����֮�����豸����������C����Թܷ����������λ����.�ڼ�G����C���ÿ3 d�������ʵ�������.�ڴ����ĵ�10��20��30 d�ֱ��C���G�����ȡ��������Ӭ���ڶ�����������DAM2��������Ӭ����Ϊ���м�⣬���������������1.3.1��ͳ�Ʒ���3 d����Ӭ���˶���˯�����.
��������ʵ���У���Ӭ���˶���˯�����ͳ�ƾ����Ը���ÿֻ��Ӭ�ľ�ֵ.���ǵ�CS��Ӭ˯����ҹ��IJ��죬���졢ҹ����һ����ҹ���ڣ�24 h�������ݷֱ����ͳ�ƣ��˶�������Ҫͨ��ͳ��Ŀ��ʱ����ڵ����˶����͵�λʱ���˶��������˶���/���ѵ�ʱ�䣩�������ۣ�˯�������Ҫͨ��ͳ��Ŀ��ʱ����ڵ�˯��ʱ�䡢˯�ߴ�����ÿ��˯��ʱ����˯��ʱ��/˯�ߴ������������ۣ���Ӭ��ֹ������ʱ�����5 minʱ���жϹ�Ӭ����˯��״̬[31].
����1.4 ����������ػ������ˮƽ�ļ��
������Զ�ģ������ЧӦ��������Ӧ�Ķ�ʱ�䴦���飬���о�����ӫ�ⶨ��PCR��һ�����ģ������ЧӦ��period(per����timeless(tim����clock(clk����cycle(cyc����cryptochrome(cry�����������������ػ�������Ӱ�죬���巽�����£�ȡ������Ӭװ���Թ��У����ô����飨G�飩�Լ������飨C�飩��G�����������λ���б�¶����3 d,C�������������ϣ�����CS��ӬΪ˫�ģ�͵ij���������[39]���ڿ��ƺ�Ϩ��ʱ����ֻ�߷�[40]�����о���6��00 pm��Ϩ��ʱ����ȡ����ÿ��10ֻ��Ӭ������EP���У�Һ���ٶ���������-80������б���.����TRIzol����ȡ������Ʒ����RNA�����ó�������ֹ��ȼƼ��RNA��Ũ�ȣ�����PrimeScript?RT Master Mix(Perfect Real Time���Լ��еIJ���ͷ�Ӧ������RNA��ת¼��cDNA������cDNA�ó���ˮϡ��15������ΪRT-PCR��ģ�壬����SYBR?Premix Ex Taq?II(Tli RNaseH Plus���IJ���ͷ�Ӧ����������Զ�����⣬RT-PCR������������TMֵΪ55�棬40��ѭ��.RT-PCR��Ӧ����rp49��Ϊ�ڲλ��������2-����Ct�������м������[41].����ʵ���ظ�3������.��������������м���1.
����Table 1 Primer sequences

����1.5�� ������ػ������ˮƽ�������ļ��
�������о���һ�������ģ������ЧӦ�Զ������ø���Ұ����ǻ�ø��ɫ�����ǻ�ø�ı������ddc��pale��trh�ı������.���л������ˮƽ�ļ�ⷽ���������ӻ����ⷽ��һ�������1.4������������������м���1.
�������⣬��һ�����ģ������ЧӦ�Զ�Ͱ���dopamine,DA����5-��ɫ����5-hydroxytryptamine,5-HT���������ʺ�����Ӱ�죬���巽�����£�ȡ������Ӭװ���Թ��У����ô����飨G�飩�Լ������飨C�飩��G�����������λ���б�¶����3 d,C��������������.��6��00 pm��Ϩ��ʱ����ȡ����ÿ��25ֻ��Ӭ������EP���У�Һ���ٶ���������-80������б��ã������Ϻ�ø���������Ͱ��Լ����Լ�����5-��ɫ���Լ��вⶨ��Ӭ����DA��5-HT�������ʵĺ���.
����1.6 ������ͳ�Ƽ�����
�������о������ݲ���Microsoft Excel��SPSS 25��������ͳ�Ʒ���.��������ö�������t�������.ͳ�ƽ������Mean±SEM��ʾ��“*”��ʾP<0.05�������飨C�飩�ʹ����飨G�飩֮��IJ���ﵽ����ˮƽ��“**”��ʾP<0.01�������飨C�飩�ʹ����飨G�飩֮��IJ���ﵽ������ˮƽ.
����2 �����
����2.1 ��ģ������ЧӦ���˶���˯����Ϊ��Ӱ��
����2.1.1 ����ʱ��ģ������ЧӦ��Ӱ��
����ѡȡ����ǰ����1 d������3 d������������1 d��5 d�����������ݽ�����Ӭ�˶������ͳ�Ʒ���.��Ӭ���˶��仯���ɱ�����C���G�����Ӭ�����ֳ�һ������ҹ�仯���ɣ���6��00 am��6��00 pm���Ҹ���һ���˶��߷��ڣ�ͼ2a�������Ӭ��������ҹ�����Լ�ʵ�������õĹ���������ͬ����Ҫ�ܹ����ڱ仯��Ӱ�죬�������ڼ������������.ͨ������ģ������ЧӦ����3 dʱ������Ӭ���˶����.�����ʾ��G����Ӭ24 h���˶�������P<0.01���͵�λʱ���˶�������P<0.05�������ӣ��������������ҹ�����˶������ʾ��G����Ӭ�������˶������͵�λʱ���˶�������C�������Ȼ�������ӣ�����δ������ˮƽ����ҹ�������˶������͵�λʱ���˶����������ӣ�P<0.01����ͼ2b��c��.ģ������ЧӦ��ǿ����Ӭҹ�����˶����͵�λʱ���˶���.
����Fig.2 Effects of simulated microgravity on the activity of Drosophila melanogaster
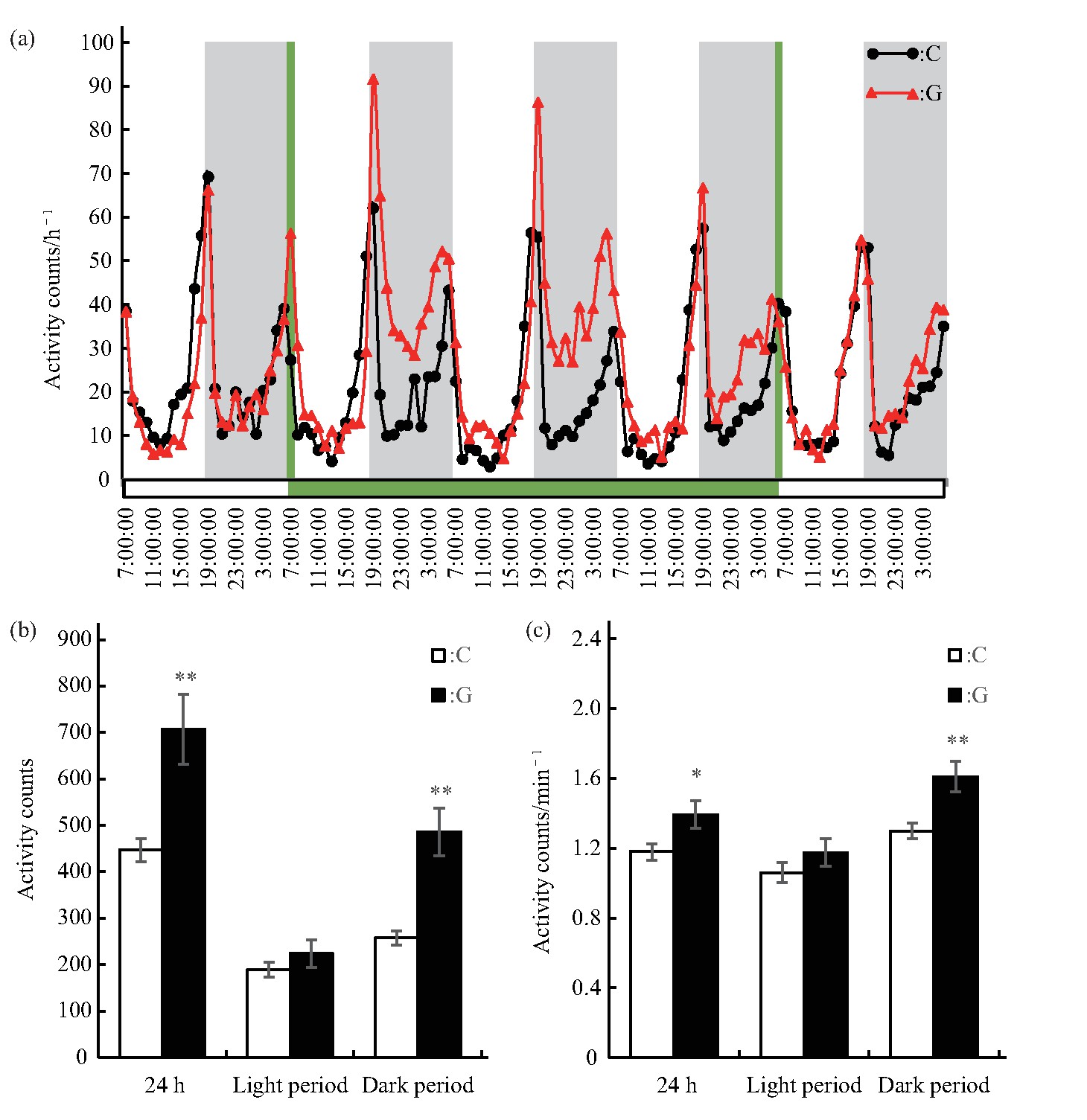
����(a)Activity rhythm,grey bars stand for nighttime,green bars stand for the time of simulated microgravity.(b)Activity counts in different periods.(c)Activity counts per minute in different periods.White and black boxes represent data of control and simulated microgravity flies,respectively.*P<0.05,**P<0.01.
������һ��������Ӭ��˯������������ʾ��C���G�����Ӭ˯�߾����ֳ�һ������ҹ�仯���ɣ���6��00 am��6��00 pm���Ҹ���һ��˯�ߵ��ڣ�ͼ3a�������Ӭ���˶��仯���ɸպ��෴��������Ҫ���������ڵı仯��أ��������������ֳ��˲���.ͬ������ͳ����ģ������ЧӦ����3 dʱ������Ӭ��˯������������ʾ��G����Ӭ24 h��˯��ʱ����٣�P<0.01����˯�ߴ������٣�P<0.01����ÿ��˯�߳������ӣ�P<0.05�����������������ҹ����˯�������ʾ��G����Ӭ����˯�ߵ�ʱ����Ȼ��C��IJ���δ������ˮƽ������˯�ߴ������٣�P<0.01����ÿ��˯�߳������ӣ�P<0.01������ʾ��Ӭ��ʱ˯�����࣬˯�������ϸߣ�ҹ��˯��ʱ����٣�P<0.01����˯�ߴ������٣�P<0.01������ÿ��˯�߳�����C��IJ���δ������ˮƽ��ͼ3b��c��d��.�����ʾ����Ӭ��˯��ʱ��ļ�����Ҫ������ҹ��˯�ߴ������٣�����˯��ʱ��ļ�����ɵ�.��ˣ���ʱ��ģ������ЧӦ����Ӭҹ����˯�ߴ������١�˯��ʱ�����̡���λʱ���˶��������ӡ��˶�������.
����Fig.3 Effects of simulated microgravity on the sleep of Drosophila melanogaster
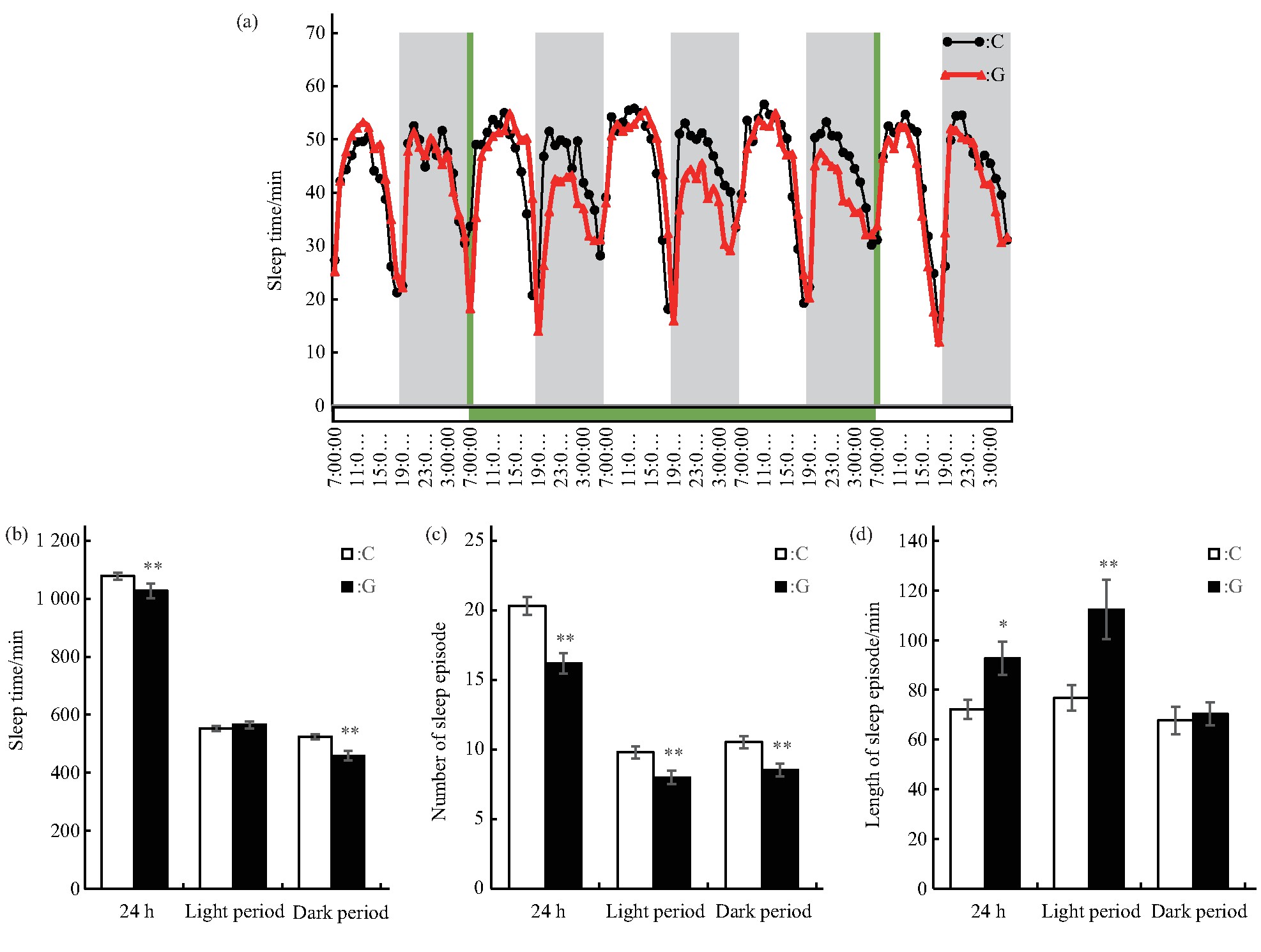
����(a)Sleep rhythm,grey bars stand for nighttime,green bars stand for the time of simulated microgravity.(b)Sleep time in different periods.(c)Sleep episode number in different periods.(d)Length of sleep episode in different periods.White and black boxes represent data of control and simulated microgravity flies,respectively.*P<0.05,**P<0.01
����2.1.2�� ��ʱ��ģ������ЧӦ��Ӱ��
�����ڶ�ʱ��ģ������ЧӦ����Ӭ�˶���˯�ߵ�Ӱ���о��У��豸�رպ���Ӭ���˶���˯����Ϊ�ܿ�ָ������յ�ˮƽ.��ˣ����о���һ�������ϳ�ʱ��ģ������ЧӦ����Ӭ�˶���˯�ߵ�Ӱ�죬�ֱ��ڴ���10 d��20 d��30 d�����ȡ����C���G�����Ӭ���ڶ��������½�������3 d����Ϊ���.��Ӭ���˶��仯���ɱ����������Ǵ���10 d��20 d����30 d,C���G�����Ӭ�����ֳ�һ������ҹ�仯���ɣ���6��00 am��6��00 pm���Ҹ���һ���˶��߷��ڣ�ͼ4a���������ʱ�䴦��һ�£���Ҫ�ܹ����ڵ�Ӱ��.��һ��������Ӭ�˶������ʾ������10 d����Ȼ��Ӭ24 h���˶�������C��IJ���δ������ˮƽ�������������˶������ӣ�P<0.05����ҹ����λʱ���˶��������٣�P<0.05��������20 d���˶������C��IJ���δ������ˮƽ������30 d��ҹ�����˶�������P<0.05���͵�λʱ���˶�������P<0.01�������٣��������C��IJ���δ������ˮƽ��ͼ4b��c).
����Fig.4 Effects of different duration of simulated microgravity on the activity of Drosophila melanogaster
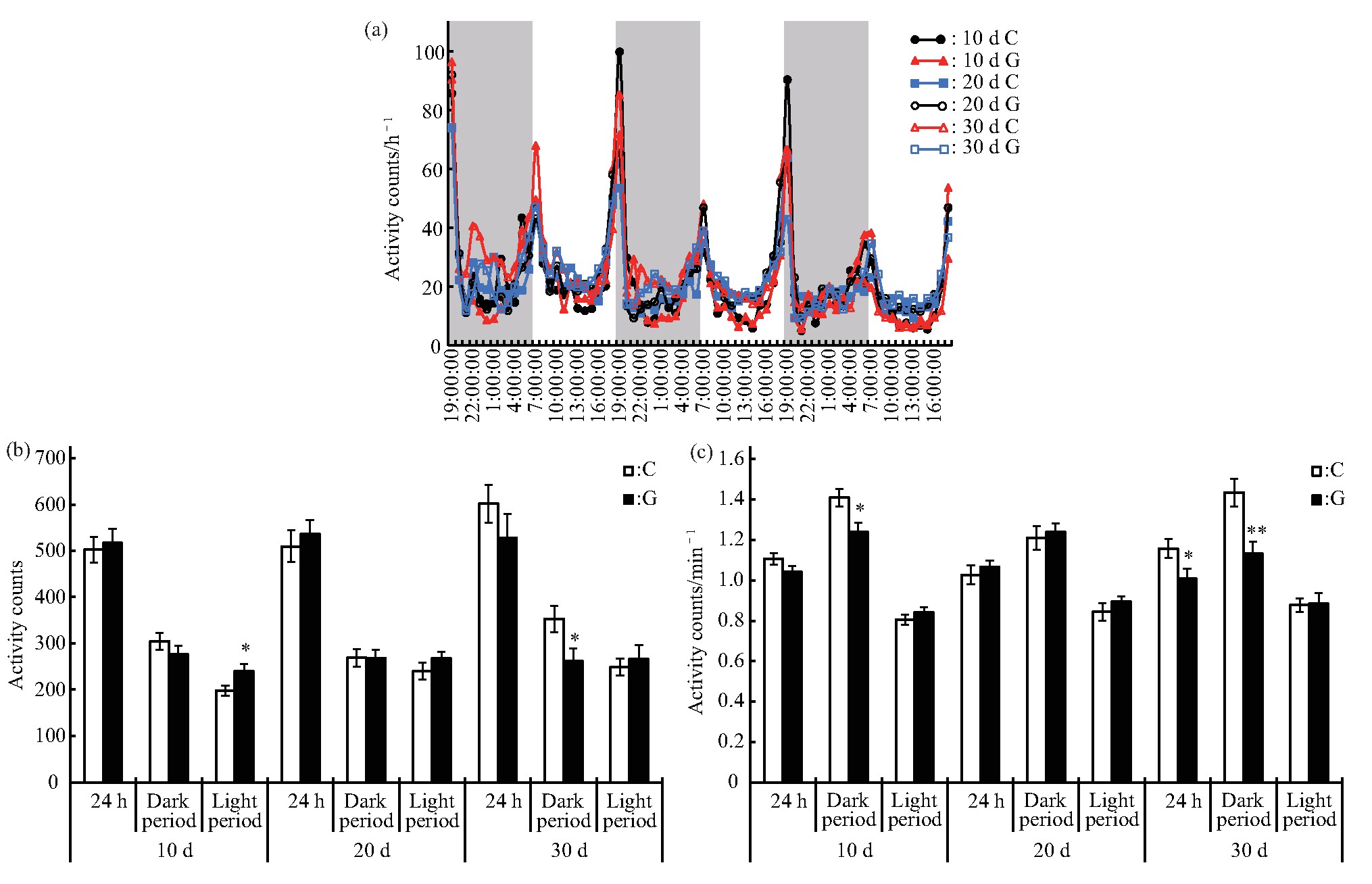
����(a)Activity rhythm after experiment,grey bars stand for nighttime.(b)Activity counts in different periods after experiment.(c)Activity counts per minute in different periods after experiment.White and black boxes represent data of control and simulated microgravity flies,respectively.*P<0.05,**P<0.01.
����˯�ߵķ��������ʾ��˯�߱仯����ͬ�����ܱ�¶ʱ���Ӱ�죬���ֳ�һ������ҹ�仯���ɣ���6��00 am��6��00 pm���Ҹ���һ��˯�ߵ��ڣ�ͼ5a�������ʱ�䴦��һ�£���Ҫ�ܹ����ڵ�Ӱ��.��һ������˯������������ʾ������10 d����Ӭ�����˯��ʱ����٣�P<0.05��������20 d��30 d���˯�߲�����C��IJ����δ������ˮƽ��ͼ5b��c��d).
����2.2 ����������ػ���ı���ˮƽ
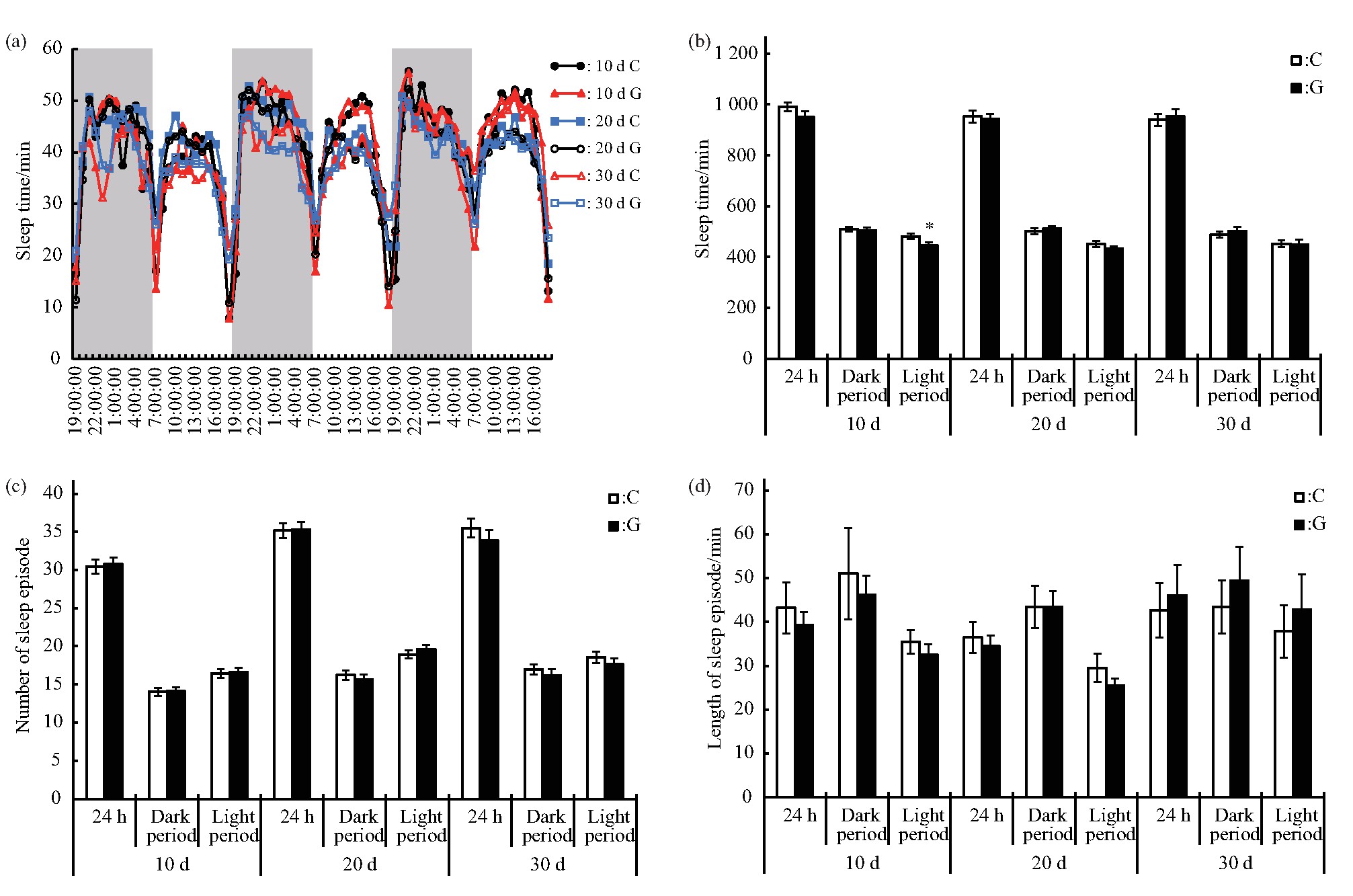
������Ӭ��˯��-����״̬�������ӵ���������أ����о�ѡȡģ������ЧӦ��¶����3 d����Ӭ����һ������˺��������ӻ���per��tim��clk��cyc��cry�ı���ˮƽ�������ʾ��per�������Ա���ˮƽ��C��IJ���δ������ˮƽ��tim��clk��cyc��cry����Ա���ˮƽ�����ӣ�P<0.01��������clk����ԼΪC���2.5����ͼ6)[42].
����Fig.5 Effects of different duration of simulated microgravity on the sleep of Drosophila melanogaster
����(a)Sleep rhythm after experiment,grey bars stand for nighttime.(b)Sleep time in different periods after experiment.(c)Sleep episode number in different periods after experiment.(d)Length of sleep episode in different periods after experiment.White and black boxes represent data of control and simulated microgravity flies,respectively.*P<0.05,**P<0.01.
����Fig.6 Effect of simulated microgravity on the relative mRNA level of selected circadian clock genes of Drosophila melanogaster
����White and black boxes represent data of control and simulated microgravity flies,respectively.*P<0.05,**P<0.01.
����2.3�� ������ػ���ı���ˮƽ������
������Ӭ��˯��-����״̬Ҳ�ܵ���ϵͳ�ĵ��ڣ����о����ģ������ЧӦ��DA��5-HT�ϳ��ж������ø�ı������ddc��DA�ϳ����Ұ����ǻ�ø�ı������pale��5-HT�ϳ���ɫ�����ǻ�ø�ı������trh����ˮƽ��Ӱ�죬�����ʾ��ddc��pale��trh�������Ա���ˮƽ�����ӣ��ֱ�ΪP<0.01,P<0.01,P<0.05����ͼ7a��.��һ���������Ӭ����DA��5-HT�ĺ����������ʾģ������ЧӦ��������ӬDA��5-HT�ĺ�����C��IJ����δ������ˮƽ��ͼ7b��c).
����Fig.7 Effects of simulated microgravity on the selected neurotransmitter of Drosophila melanogaster
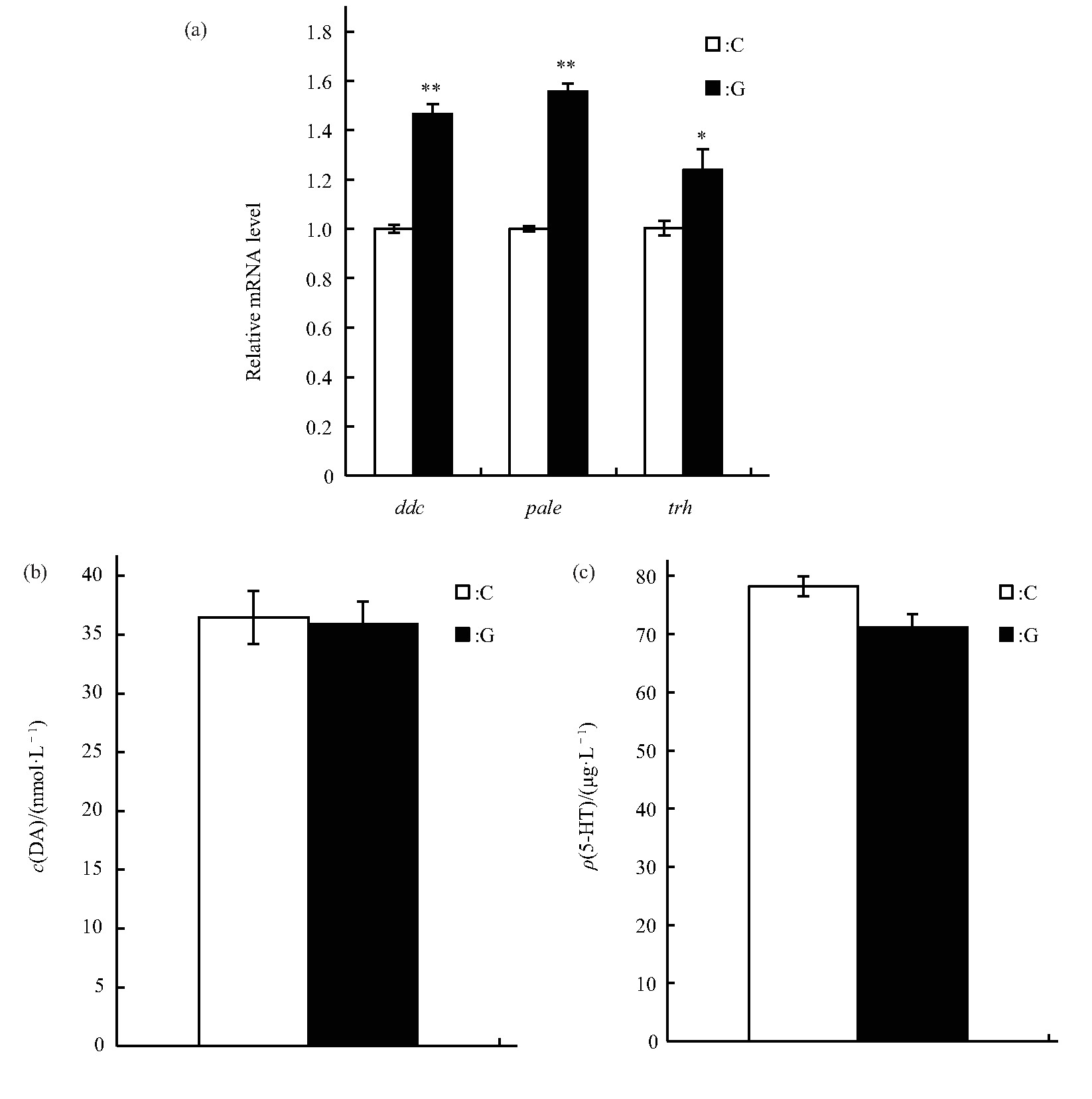
����(a)Relative mRNA level.(b)Content of DA.(c)Content of 5-HT.White and black boxes represent data of control and simulated microgravity flies,respectively.*P<0.05,**P<0.01.
����3 ������
���������ҹ�����Ա���칤��ʦδ���ڹ칤��ʱ����ӳ�����α��Ϻ���Ա�ڹ��ڼ�ĸ�����˯�������ڹ콡����̬��ά���Ѿ���ΪӰ���Һ�����ҵ��Զ��չؽ���������Ҫ�ؼ�����֮һ.��Ŀǰ���ڿռ价����˯�ߵ�Ӱ�켰���û����˽�����.�����о����������е������λ�ǿ�չģ������ЧӦ�Թ�Ӭ˯����Ϊ�о��Ŀ����ԣ�������̽��ģ������ЧӦ�Թ�Ӭ�˶���˯�ߵ�Ӱ��.
������ʱ��ģ������ЧӦ���������������Ӭ�˶���˯�߽��ɵı仯�����������й����ڵı仯һ��.һ���棬������ģ������ЧӦ���˶���˯�߽���û��Ӱ�죻��һ���棬���ǵ�����ǿ��ʱ���ӣ�ģ������ЧӦ��Ӱ����ܽ�������������һ���̶�����Ӱ��ᱻ���������ڸ�[31].�ý��Ҳ�Ӳ���˵�������λ��������ת��������Ӱ���Ӭ�Ĺ��н���.�Ӷ�ʱ���ģ������ЧӦ��������Ӭ�˶���˯�ߵķ���������Կ�����ģ������ЧӦ��ǿ��ҹ����Ӭ���˶�����������Ҫ��ӳ����Ӭҹ��˯��ʱ���˯�ߴ����ļ��٣�����ƽ��ÿ��˯��ʱ�����ӣ���������Ӭҹ�����˶�������˯��ʱ����ٶ�������˯�߲����������[43,44,45,46]�����켰ҹ����˯�ߴ��������٣����ܱ�����Ӭ��ʵ�鴦���¸�����˯��������ҹ��.Nike-Orion̽�ջ���ϴ��صĹ�Ӭ�����������ڼ䣬Ҳ�۲쵽�����˶�����ֵ������ӵ����[32]�����⣬��IML-2����̫�������У�������¶�ڼ䣬��Ӭ���˶�����[47]���������߷�����Ҳ�����Ƶķ���[48]���뱾�о��Ľ��һ��.ǰ������Ա�ķ��б���ͼ����[4]Ҳ����������Ա�ڹ��ڼ�˯��ʱ����١�˯�����������뱾�о�����Ӭ˯��ʱ����ٵĽ��һ��.�ڱ��о��У���Ӭ˯��ʱ��ļ�����Ҫ��˯�ߴ������ٵ��µģ�����ҪӰ�����ҹ��˯��.
������ʱ��ģ������ЧӦ�����������������30 d����Ӭҹ�����˶��������Ʋ��뼡������[49,50,51,52]���ӿ�˥��[20,47,53]����أ���IML-2����̫�������У���Ϊ��14.5 d�ĺ������֮����Ӭ��ȡ�»ָ�����ֳ������ϻ���Ӧ[47].����Ҳ��ģ������ЧӦ����Ӭ�������������о�����û�з���������Ӱ�죨��¼S1��ͼS2�������ǵ��ռ价���ĸ����ԣ����������Ƿ�����Эͬ���ã�ʹ�ú���ɻ���¶�ı仯��Ϊ���ԣ������һ���о�.�������⣬����Ӭ�˶���˯�ߵ�����Ӱ���С������Ҫ��������ԭ��.һ���棬���Ŵ���ʱ����ӳ�����Ӭ��������Ӧ�ԣ�Nike-Orion̽�ջ���ϴ��صĹ�Ӭ��������˲̬��Ӧ�������˶�ˮƽ�������ĵ�2 min�ﵽ���壬֮��������½�����������ʱ��ֻ��3.8 min�����ó���ʱ��Ӱ��[32]�������о������������������߷����У������Ӭ�ڵ�һ�ε�0 g���˶����ӣ��������θ�ЧӦ����[48]������ʱ����еĺ���Ա˯������ĵ�����Ҳ����ͬ������������ֺ���Ա�ڷ��й����е�ǰ30 d˯��ʱ�����ż��٣������ڻ���һ���ĺ�ת[54].��һ���棬���ڳ�ʱ�䴦������Ӭ�Ƿ����ڶ��ջ����н��м�⣬��˸ý��Ҳ��ģ������ЧӦ�������Ӭ�ָ���أ��Ӷ�ʱ��Ĵ�����Ҳ���������Ƶ���������豸�رպ���Ӭ���˶���˯�߶��ָ������������.��ˣ�������Ϊ������������к�ǿ����Ӧ�������Ҳ����IJ���Σ���ǿɻָ��ģ��Ӻ���Ա�ص�������˯������о�Ҳ�������Ƶ����[4].
����Ϊ�˽�һ������ģ������ЧӦӰ���Ӭ˯�ߵĻ��ƣ����о��ӹ�Ӭ˯�ߵ��ص������Ӻ����ʽ��з���.�����ӵ��ػ��������ּ�߶ȱ���[55,56,57,58]��������ת¼-����ĸ�������·���ƣ�clk��cycת¼������γɵ�������嵰��CLK-CYC�ٽ�per��tim����ı����per��timת¼������γ�������嵰��PER-TIM����������CLK-CYC[33,57,59,60,61]����CLK����Ϊ���������Ƽ�[62,63].�����ֽ��ɵ���ʼ�����λ���Ա����ձ�¶�������ã����ܵ���Ϊcry�����CRY������Ϊ�����壬��TIM�����������Խ�ϣ�ʹPER���ȶ����Ӷ����ٶ�CLK-CYC������[64].���о����֣���ʱ���ģ������ЧӦ������tim��clk��cyc��cry�������Ա���ˮƽ�����ӣ���˵���������ܹ���ģ������ЧӦ������Ӧ.���б������ڹ���к���յĹ�Ӭ����ת¼����������ֺ��������ӣ�per��tim��cry����û�����Ա仯[31]�����������ʵ�����ڷ��н���48 h����е���أ��ɱ��о�ǰ������ɼ���Ӭ�к�ǿ�Ļָ���������������Թ�Ӭ��Ӱ������Ѿ��ָ�.������ϵͳ���棬���о��Զ�ʱ�䴦�����������ر������ı���ˮƽ���Լ����ʺ��������˲ⶨ��ѡ��������DA��5-HT,DA�ܹ��ٽ����ѣ���5-HT�����������͵IJ�ͬ�ٽ����ѻ�˯��[65,66,67,68].�������ģ������ЧӦ������ddc��pale��trh����Ա���ˮƽ�����ӣ�˵��ģ������ЧӦ������ǿ��������ʵĺϳɣ����ǽ�һ���ļ����ȴ����DA��5-TH�ĺ�����û�г��ֱ仯��������뱾�о���ⷽʽ���.���о��������Ӭ�����DA��5-TH����������˯�ߵ�����Ҫ���Բ�������ϸ��������ʣ����������ܵ����ʺ���������ȶ�.ͬʱ����ϵͳ�ĵ��ڲ����ǿ���һ�����ʻ��أ�����������ϵͳ���ӵĹ�ͬ����[65].
������Ȼ�����λ�Dz������ṩ�������������������ӹ�Ӭ�˶���˯�߽��ɼ���ز����ı仯�����������Ϊ���������λ��ģ������ЧӦ��¶�����£��ھ��ýΣ���Ӭ������Ͷ�������˶����ߺ�˯�����߸߶��غϣ���������ЧӦģ��Σ���Ӭ��Ȼ���ֳ�����еĽ�����Ϊ.ͨ�����۹۲�Ҳ�������豸ת�������У���Ӭ����磬��������˦������ת�����е���Ϊ���֣���¼S1��ͼS1��.ͬʱ������Ҳ����Ӭ����Ӧ�����ø���Ժͻ�������˼�⣬Ҳδ����������Ӱ�죨��¼S1��ͼS4��S5��.���о����֣�ģ������ЧӦ�£���ʱ������Ӭҹ�����˶�������ǿ��˯���٣�����������˯�߲�����������ģ������ЧӦ�Ľ�����ӳ�����ЩӰ�춼�ܵõ��ָ�����Щ���Ӭ�����Ӻ����ʵĵ�����أ�����ػ��ƴ���һ���о�.
������ˣ����о���Ϊ����ģ������ЧӦ�о������Թ�Ӭ�˶���˯�ߵ�Ӱ�����һ���Ŀ����ԣ�����о�����ܹ�Ϊ����ҽѧ�о��������Ǻ��컷���Ժ���Ա˯�ߵ�Ӱ���о��ṩһ���IJο�������Ҫע����ǣ����������λ��ģ������ЧӦȷʵ����һ���IJ��㣬�ر�������Ϊ�о�����Ӧ�ü��٣�����ϣ����һЩ�������о���ͬʱҲ�ڻ���̽�����õķ�������Ŭ����չһЩ�ڹ�IJ��ԣ�ͨ����-�ضԱ�ʵ��ļ��飬�ܹ����õ���֤�ػ�ʵ�����ؽ����Ϊ���۸÷����Ŀ����ԺͿ�ѧ���ṩ����IJο�����.
���������
����[1] Itani O, Jike M, Watanabe N, et al. Short sleep duration and health outcomes:a systematic review, meta-analysis, and metaregression. Sleep Med, 2017, 32:246-256
����[2] Afolalu E F, Ramlee F, Tang N K Y. Effects of sleep changes on pain-related health outcomes in the general population:a systematic review of longitudinal studies with exploratory metaanalysis. Sleep Med Rev, 2018, 39:82-97
����[3] Khan S, Duan P F, Yao L G, et al. Shiftwork-mediated disruptions of circadian rhythms and sleep homeostasis cause serious health problems. Int J Genomics, 2018,2018:8576890
����[4] Barger L K, Flynn-Evans E E, Kubey A, et al. Prevalence of sleep deficiency and use of hypnotic drugs in astronauts before, during,and after spaceflight:an observational study. Lancet Neurology,2014, 13(9):904-912
����[5]����·,��ѧ��,�غ���,��.���˺���˯�����⼰���Ԥ��ʩ.�վ�ҽѧ��־,2014,30(01):57-60Jing X L, Liu X Y, Qing H B, et al. Medical Journal of Air Force,2014, 30(01):57-60
����[6] Brainard G C, Barger L K, Soler R R, et al. The development of lighting countermeasures for sleep disruption and circadian misalignment during spaceflight. Curr Opin Pulm Med, 2016,22(6):535-544
����[7] Caddick Z A, Gregory K, Flynn-Evans E E. Sleep environment recommendations for future spaceflight vehicles[M]//STANTON N A, LANDRY S, DIBUCCHIANICO G, et al. Advances in Human Aspects of Transportation. Cham; Springer International PublishingAg. 2017:923-933
����[8]��һ��,�DZ�,��ӱ.����Ա�������˶�ģ��ѵ���������о�����.���˺���,2018,24(02):227-237Jiang Y F, Qiao B, Zhao Y. Manned Spaceflight, 2018, 24(02):227-237
����[9] Frey D J, Ortega J D, Wiseman C, et al. Influence of zolpidem and sleep inertia on balance and cognition during nighttime awakening:a randomized placebo-controlled trial. J Am Geriatr Soc, 2011, 59(1):73-81
����[10] Hoque R, Chesson A L, Jr. Zolpidem-induced sleepwalking, sleep related eating disorder, and sleep-driving:fluorine-18-flourodeoxyglucose positron emission tomograph analysis, and a literature review of other unexpected clinical effects zolpidem.Journal of Clinical Sleep Medicine, 2009, 5(5):471-476
����[11] Van Oosterhout W P J, Terwindt G M, Vein A A, et al. Space headache on Earth:head-down-tilted bed rest studies simulating outer-space microgravity. Cephalalgia, 2015, 35(4):335-343
����[12] Watenpaugh D E. Analogs of microgravity:head-down tilt and water immersion. Journal of Applied Physiology, 2016, 120(8):904-914
����[13] Wang C Y, Wu J H, Cui Y, et al. Simulated weightlessness by tailsuspension affects follicle development and reproductive capacity in rats. Int J Clin Exp Pathol, 2016, 9(12):12208-12218
����[14] Sung M, Li J, Spieker A J, et al. Spaceflight and hind limb unloading induce similar changes in electrical impedance characteristics of mouse gastrocnemius muscle. J Musculoskelet Neuronal Interact, 2013, 13(4):405-411
����[15] Van Loon J J W A. Some history and use of the random positioning machine, RPM, in gravity related research. Advances in Space Research, 2007, 39(7):1161-1165
����[16] BorstAG, Van Loon J J WA. Technology and developments for the random positioning machine, RPM. Microgravity Science and Technology, 2008, 21(4):287-292
����[17] Wuest S L, Richard S, Kopp S, et al. Simulated microgravity:critical review on the use of random positioning machines for mammalian cell culture. Biomed Research International, 2015,2015:971474
����[18] Brungs S, Egli M, Wuest S L, et al. Facilities for simulation of microgravity in the ESA ground-based facility programme.Microgravity Science and Technology, 2016, 28(3):191-203
����[19]��ܿ.��ͬ�����Թ�Ӭ�ߺ����ٶȼ���ػ����Ӱ��[D].����:�������´�ѧ,2012Wu Y. The Effects of Drosophila Development Rate After Embryo and Related Genes by Different Gravities[D]. Dalian:Dalian Maritime University, 2012
����[20]���˾�.ģ������ЧӦ�Թ�Ӭ��������ػ����Ӱ��[D].����:�������´�ѧ,2010Weng Y J. Simulated Microgravity Effects on Drosophila Development and Related Genes[D]. Dalian:Dalian Maritime University, 2010
����[21]��Զ��,��ά��,��³��,��.�������ͻ�ת��.����ҽѧ��ҽѧ����,2008,4:368-371Jiang Y D, Li W N, Wang L F, et al. Space Medicine&Medical Engineering, 2008, 4:368-371
����[22]��С��,��־��,������,��.�����в��ܳ���֮��——����������������ҹ���ɵı仯�о�.������ѧ, 2015, 27(11):1433-1440Liang X D, Liu Z Z, Chen X Y, et al. Chinese Bulletin of Life Sciences, 2015, 27(11):1433-1440
����[23] Schwarzenberg M, Pippia P, Meloni M A, et al. Signal transduction in T lymphocytes-A comparison of the data from space, the free fall machine and the random positioning machine[M]//HASENSTEIN K H, SATO A, COGOLI A, et al. Life Sciences:Microgravity Research Ii. 1999:793-800
����[24] Briegleb W. Some qualitative and quantitative aspects of the fastrotating clinostat as a research tool. ASGSB bulletin:publication of the American Society for Gravitational and Space Biology,1992, 5(2):23-30
����[25] Hoson T, Kamisaka S, Masuda Y, et al. Evaluation of the threedimensional clinostat as a simulator of weightlessness. Planta,1997, 203 Suppl:S187-S197
����[26]����.����ڵ������ģ��ռ�����������ЧӦ:�ӿռ�ϸ��������������Ӧ̸��.��ѧͨ��,2014,59(20):2004-2015Long M. Chin Sci Bull(Chin Ver), 2014, 59(20):2004-2015
����[27] Herranz R, Benguria A, Lavan D A, et al. Spaceflight-related suboptimal conditions can accentuate the altered gravity response of Drosophila transcriptome. Mol Ecol, 2010, 19(19):4255-4264
����[28] Herranz R, Lavan D A, Medina F, et al. Drosophila GENE experiment in the Spanish soyuz mission to the ISS:II. Effects of the containment constraints. Microgravity Science and Technology, 2009, 21(4):299-304
����[29] Marcu O, Lera M P, Sanchez M E, et al. Innate immune responses of Drosophila melanogaster are altered by spaceflight. Plos One,2011, 6(1):e15361
����[30] Taylor K, Kleinhesselink K, George M D, et al. Toll mediated infection response is altered by gravity and spaceflight in Drosophila. Plos One, 2014, 9(1):e86485
����[31] Ma L L, Ma J, Xu K Y. Effect of spaceflight on the circadian rhythm, lifespan and gene expression of Drosophila melanogaster.Plos One, 2015, 10(3):e0121600
����[32] Miller M S, Keller T S. Drosophila melanogaster(fruit fly)locomotion during a sounding rocket flight. Acta Astronautica,2008, 62(10-11):605-616
����[33] Vosshall L B, Price J L, Sehgal A, et al. Block in nuclearlocalization of period of protein by a 2nd clock mutation, timeless.Science, 1994, 263(5153):1606-1609
����[34] Smith R F, Konopka R J. Effects of dosage alterations at the per locus on the period of the circadian clock of Drosophila. Molecular&General Genetics, 1982, 185(1):30-36
����[35]��˼��.���ڹ�Ӭģʽ�����˯�����������Ե��о�[D];��������ҽҩ��ѧ,2014Liu S Q. Research of sleep and genetic correlation based on drosophila's biological model; Heilongjiang University Of Chinese Medicine, 2014
����[36]������,����,������.�����ӵ�����——2017��ŵ��������ѧ��ҽѧ�����.��ҽ�ƴ�ѧѧ��,2017,38(05):765-769Li Y Z, Zhang T, Wang X M. Journal of Capital Medical University, 2017, 38(05):765-769
����[37] Guo F, Holla M, Diaz M M, et al. A circadian output circuit controls sleep-wake arousal in Drosophila. Neuron, 2018, 100(3):624-635
����[38] Harbison S T, Kumar S, Huang W, et al. Genome-wide association study of circadian behavior in Drosophila melanogaster. Behav Genet, 2019, 49(1):60-82
����[39]����.������Ϊѧ����[M]. 1 ed.����:��ѧ������, 2009Qing Y C. Introduction to insect behavior. Beijing:Science Press,2009
����[40] Dubowy C, Sehgal A. Circadian rhythms and sleep in Drosophila melanogaster. Genetics, 2017, 205(4):1373-1397
����[41] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C)method. Methods, 2001, 25(4):402-408
����[42] Zhang H, Liao Y, Zhang Z, et al. Effect of simulated microgravity on the circadian rhythm of Drosophila melanogaster. New Biotechnology, 2018, 44(Suppl):S83
����[43] Hendricks J C, Finn S M, Panckeri K A, et al. Rest in Drosophila is a sleep-like state. Neuron, 2000, 25(1):129-138
����[44] Huber R, Hill S L, Holladay C, et al. Sleep homeostasis in Drosophila melanogaster. Sleep, 2004, 27(4):628-639
����[45] Sitaraman D, Aso Y, Jin X, et al. Propagation of homeostatic sleep signals by segregated synaptic microcircuits of the Drosophila mushroom body. Curr Biol, 2015, 25(22):2915-2927
����[46] Akerstedt T, Kecklund G, Ingre M, et al. Sleep homeostasis during repeated sleep restriction and recovery:support from EEG dynamics. Sleep, 2009, 32(2):217-222
����[47] Benguria A, Grande E, Dejuan E, et al. Microgravity effects on Drosophila melanogaster behavior and aging. Implications of the IML-2 experiment. Journal of Biotechnology, 1996, 47(2-3):191-201
����[48] Lebourg E, Grimal A, Fresquet N, et al. Spontaneous locomotoractivity of Drosophila melanogaster flies at various gravity levels(0 G, 1 G, 1.8 G)during parabolic flights. Behavioural Processes,1995, 34(2):175-184
����[49] Guo N, Fan X Y, Wu Y T, et al. Effect of constraint loading on the lower limb muscle forces in weightless treadmill exercise. J Healthc Eng, 2018, 2018:8487308
����[50] Nikawa T, Ishidoh K, Hirasaka K, et al. Skeletal muscle gene expression in space-flown rats. Faseb J, 2004, 18(3):522-524
����[51]������,��ܲ��,�ᄅ.ʧ��״̬��ɼ�ή�����о����չ.�й���֯�����о�,2013,17(28):5249-5254Han Z Y, Jia Y J, Tian J. Chinese Journal of Tissue Engineering Research, 2013, 17(28):5249-5254
����[52]������,������,����÷,��.�����������彡��ЧӦ�о���չ.����ҽѧ,2018,42(04):317-321Sun Y Y, Zhang Z Y, Huang X M, et al. Mil Med Sci, 2018, 42(04):317-321
����[53] Vernikos J, Schneider V S. Space, gravity and the physiology of aging:parallel or convergent disciplines? A mini-review.Gerontology, 2010, 56(2):157-166
����[54] Gundel A, Drescher J, Polyakov V V J H F, et al. Quantity and quality of sleep during the record manned space flight of 438 days.Human Factors andAerospace Safety, 2001, 1(1):87-98
����[55] Hardin P E. Transcription regulation within the circadian clock:the E-box and beyond. J Biol Rhythms, 2004, 19(5):348-360
����[56] Hardin P E. Molecular Genetic analysis of circadian timekeeping in Drosophila[M]//BRODY S. Genetics of Circadian Rhythms.San Diego; ElsevierAcademic Press Inc. 2011:141-173
����[57] Allada R. Circadian clocks:a tale of two feedback loops. Cell,2003, 112(3):284-286
����[58] Brown S A, Kowalska E, Dalmann R.(Re)inventing the circadian feedback loop. Developmental Cell, 2012, 22(3):477-487
����[59] Hardin P E, Hall J C, Rosbash M. Feedback of the Drosophila period gene-product on circadian cycling of its messenger-RNA levels. Nature, 1990, 343(6258):536-540
����[60]������,��ɭ.�����ӷ�����������.�����Ļ�ѧ,2016, 36(03):390-396Yang LT, Liu S. Chemistry of Life, 2016, 36(03):390-396
����[61]���,�ڶ�,����,��.�����ӻ����о��½�չ.�Ŵ�, 2004,26(01):89-96Li J C, Yu D, Wang F, et al. Hereditas(Beijing), 2004, 26(01):89-96
����[62]��ǿ.������:����ͻ�����������Ϊ——2017��ŵ��������ѧ��ҽѧ�����.��Ȼ��־,2017,39(06):411-416Yu Q. Chinese Journal of Nature, 2017, 39(06):411-416
����[63] Darlington T K, Wager-Smith K, Ceriani M F, et al. Closing the circadian loop:CLOCK-induced transcription of its own inhibitors per and tim. Science, 1998, 280(5369):1599-1603
����[64] Myers M P, Wagersmith K, Rothenfluhhilfiker A, et al. Lightinduced degradation of TIMELESS and entrainment of the Drosophila circadian clock. Science, 1996, 271(5256):1736-1740
����[65] Holst S C, Landolt H-P. Sleep-wake neurochemistry. Sleep Medicine Clinics, 2018, 13(2):137-146
����[66]���,Ƚ����,ŷ������,��.����ɫ����˯��-����������.�ִ�����ҽѧ��չ,2015,15(11):2191-2194Yang C, Ran M Z, Ouyang P R, et al. Progress in Modern Biomedicine, 2015, 15(11):2191-2194
����[67] Monti J M, Jantos H. The role of serotonin 5-HT7 receptor in regulating sleep and wakefulness. Rev Neurosci, 2014, 25(3):429-437
����[68] Qian Y, Cao Y, Deng B, et al. Sleep homeostasis regulated by5HT2b receptor in a small subset of neurons in the dorsal fanshaped body of drosophila. Elife, 2017, 6:e26519





