摘要:病毒病是危害西番莲产业的主要病害之一, 严重影响西番莲的产量和品质。目前, 国外对西番莲病毒病研究较多, 并取得了一定进展。中国西番莲病毒相关研究仅涉及病原鉴定, 在病毒流行病学、致病机理和抗病资源鉴选等方面鲜有报道。本文重点综述了国内外已经报道的西番莲病毒种类、侵染特性、检测技术以及主要防治策略, 并对西番莲病毒病的主要研究方向进行了展望, 以期为中国西番莲病毒病的研究和防治提供参考。
关键词:西番莲; 病毒病; 病毒检测; 防治;
Research Progress on Viruses of Passionfruit
Abstract:Virus diseases are considered as one of the most important diseases of passionfruit, and affected the yield and quality of passion fruit seriously. At present, there were many studies about virus diseases of passionfruit in some other countries, and significant progress had been made. In China the research was related to the pathogen identification of passion fruit, but there were few studies about virus epidemiology, pathogenesis and selection of disease resistance germplasm of passionfruit. The virus diseases infecting the passionfruit and their biological characteristics, pathogenic symptoms, detection and main control strategies in China and other countries were reviewed, and a prospect for research interest of virus diseases infecting passionfruit was made in order to provide references for the research and control of virus diseases infecting passionfruit in China.
Keyword:passionfruit; viral disease; virus detection; control;
西番莲属 (Passiflora L.) 是西番莲科 (Passifloracea) 中最大的一个属, 约有530个种和400多个人工杂种, 其中可食用的有80余种[1-2].因其果汁中含有多种水果的香味成分, 故又称之为百香果[3].西番莲集食用、药用和观赏价值于一身, 具有重要的经济意义[3].中国台湾于1901年从日本引入西番莲, 而后传至大陆, 至今已在台湾、广东、海南、福建、贵州、广西、云南、四川等省区推广种植[4-5].由于西番莲多为引种栽培, 各地区之间苗木销售活动频繁, 加速了病害的传播和蔓延。目前已经报道的西番莲病害主要有病毒病、细菌性病害、真菌性病害以及线虫引起的病害[6].
病毒病潜伏期长、对西番莲产业危害较大。台湾在1982年曾是亚洲最大的西番莲产区, 种植面积达1 392 hm2, 因病毒病肆虐, 导致种植面积大为缩减, 至1988年, 种植面积不足原来的一半[7].目前, 国内外已经报道了26种侵染西番莲的植物病毒, 在中国发现的有黄瓜花叶病毒、东亚西番莲病毒、西番莲斑驳病毒、广东番木瓜曲叶病毒、大戟曲叶病毒和夜来香花叶病毒等[8-11].本文重点综述了国内外已经报道的西番莲病毒的种类、侵染特性和检测技术, 以引起西番莲木质化病的豇豆蚜传花叶病毒为例, 介绍了西番莲病毒病的主要防治策略, 并对西番莲病毒病的主要研究方向进行了展望。
1、西番莲病毒病种类及侵染特性
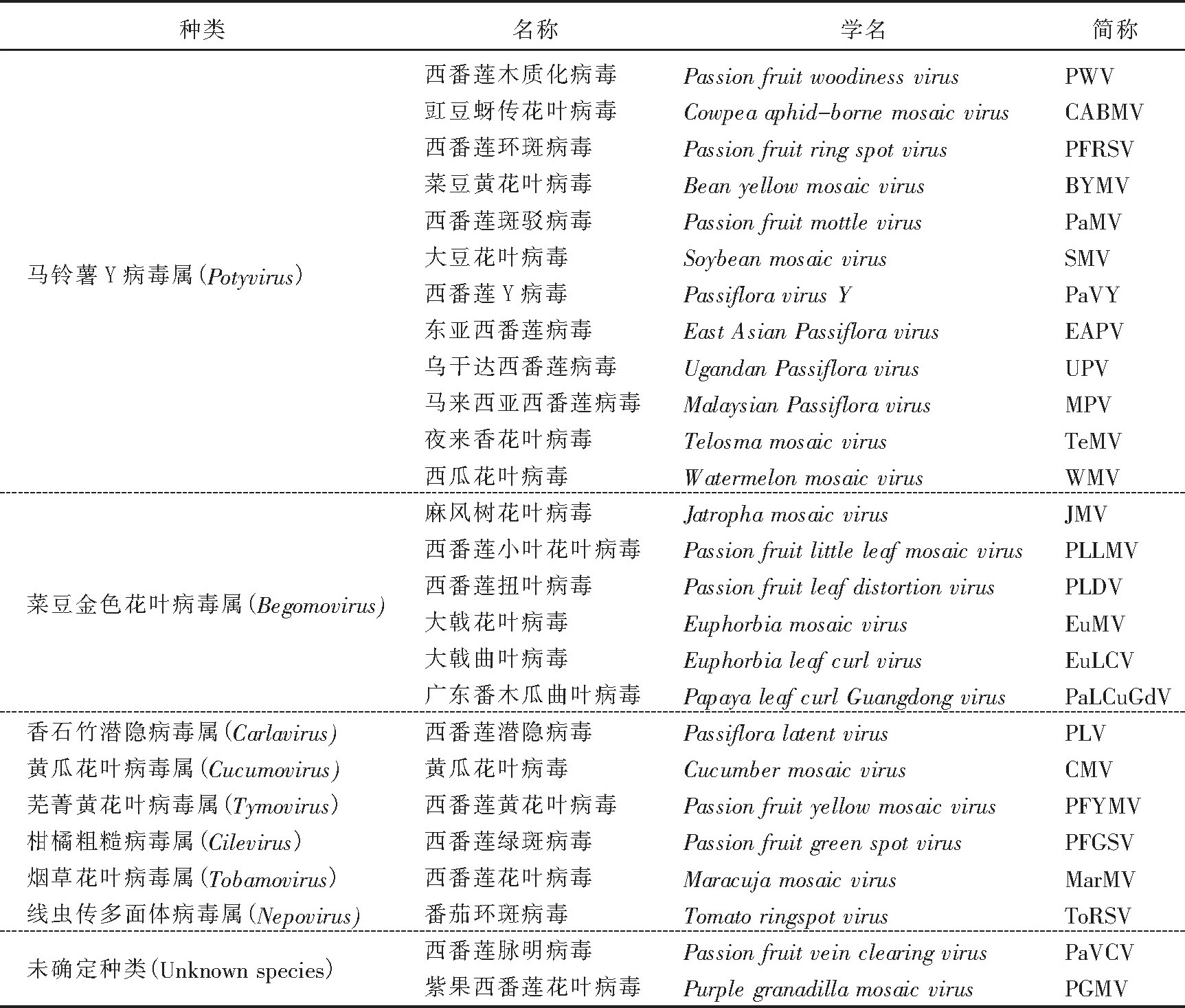
1948年, 澳大利亚报道了首例侵染西番莲的植物病毒。迄今为止, 世界范围内已发现26种能侵染西番莲的植物病毒, 包括马铃薯Y病毒属 (Potyvirus) 、菜豆金色花叶病毒属 (Begomovirus) 、香石竹潜隐病毒属 (Carlavirus) 、黄瓜花叶病毒属 (Cucumovirus) 、芜菁黄花叶病毒属 (Tymovirus) 、柑橘粗糙病毒属 (Cilevirus) 、烟草花叶病毒属 (Tobamovirus) 、线虫传多面体病毒属 (Nepovirus) 以及2种分类尚不明确的病毒 (表1) .
1.1 马铃薯Y病毒属
马铃薯Y病毒属 (Potyvirus, PVY) 是马铃薯Y病毒科 (Potyviridae) 成员, 根据寄主植物症状反应和血清学反应类型可划分为200多个株系[12].西番莲木质化病毒 (Passion fruit woodiness virus, PWV) 属于PVY, 最初发现于澳大利亚[13], 随后在尼日利亚[14]、日本[15]均有相关报道, 近年来在澳大利亚又发现了2个新的毒系, PWV-MU2和PWV-Gld[16-17].此外, 侵染西番莲的PVY还包括豇豆蚜传花叶病毒 (Cowpea aphid-borne mosaic virus, CABMV) [18-19]、西番莲环斑病毒 (Passionfruit ringspot virus, PFRSV) [20]、菜豆黄花叶病毒 (Bean yellow mosaic virus, BYMV) [21-22]、西番莲斑驳病毒 (Passionfruit mottle virus, Pa MV) [9]、大豆花叶病毒 (Soybean mosaic virus, SMV) [23]、西番莲Y病毒 (Passifloravirus Y, Pa VY) [24]、东亚西番莲病毒 (East Asian Passiflora virus, EAPV) [25]、乌干达西番莲病毒 (Uganda Passiflora virus, UPV) [26]、马来西亚西番莲病毒 (Malaysian Passiflora virus, MPV) [27]、夜来香花叶病毒 (Telosma mosaic virus, Te MV) [28]和西瓜花叶病毒 (Watermelon mosaic virus, WMV) [29].马铃薯Y病毒粒子呈线状, 无包膜, 长度680~900 nm, 直径12~15 nm, 线形正义单链RNA约含10 000个核苷酸, 编码的多聚蛋白可裂解为多个蛋白质[12].
上述病毒虽然都属于PVY, 但侵染西番莲引起的症状却存在差异。PWV和CABMV导致严重的花叶、叶片皱缩、畸形, 抑制植株生长发育, 导致果实木质化和畸形[6];SMV引起西番莲花叶、落叶, 严重时导致植株过早衰亡[23];PFRSV侵染表现为嫩叶上有黄色斑纹、环斑, 甚至产生严重的花叶和畸形叶, 而果实上无明显症状[6];Pa MV和Pa VY引起的症状相似, 叶片上呈现轻微斑纹, 而Pa MV还导致果皮上出现斑纹[6];EAPV侵染引起叶片上形成褪绿斑, 同时造成果皮褪色或出现斑纹[25];Te MV侵染引起花叶、叶片皱褶和畸形[28];WMV主要侵染紫果西番莲, 导致花叶和严重的叶片皱缩[29].PVY主要通过桃蚜 (Myzus persicae) 、棉蚜 (Aphis gossypii) 、绣线菊蚜 (Aphis spiraecola) 和橘蚜 (Toxoptera citricidus) 以非持久性方式传播[6].
1.2 菜豆金色花叶病毒属
菜豆金色花叶病毒属 (Begomovirus) 是双生病毒科 (Geminiviridae) 成员, 在波多黎各[30-31]、巴西[32]、美国[33-34]和中国[10]均有关于其侵染西番莲的报道。在波多黎各, 西番莲菜豆金色花叶病毒最初命名为西番莲叶斑驳病毒 (Passifloraleaf mottle virus, PFLMV) [30], 随后被鉴定为麻风树花叶病毒 (Jatropha mosaic virus, JMV) [31].巴西西番莲菜豆金色花叶病毒被命名为西番莲小叶花叶病毒 (Passion fruit little leaf mosaic virus, PLLMV) [32].美国西番莲菜豆金色花叶病毒有2个分离物, 分别是西番莲扭叶病毒 (passion fruit leaf distortion virus, PLDV) [33]和大戟花叶病毒 (Euphorbia mosaic virus, Eu MV) [34].在中国报道的菜豆金色花叶病毒则为大戟曲叶病毒 (Euphorbia leaf curl virus, Eu LCV) 和广东番木瓜曲叶病毒 (Papaya leaf curl Guangdong virus, Pa L-Cu Gd V) [10].
JMV侵染将引起叶片严重卷曲、变形;果实和叶片上均出现斑纹, 导致品质和产量下降[31].研究表明, 这种病毒可以从西番莲传染到菜豆, 从菜豆传染到菜豆, 但不能从菜豆传染到西番莲, 不能通过汁液摩擦传播, 也不会随带毒树的种子传播, 主要通过B型烟粉虱 (Bemisia tabaci) 传播[31].PLLMV引起严重的黄色花叶, 叶片迅速脱落, 严重影响植株生长发育, 树体结果少且多数果实畸形[31].PLLMV能通过嫁接途径侵染翅茎西番莲 (Passiflora alata) 、大果西番莲 (Passiflora quadrangularis) 和细柱西番莲 (Passiflora suberosa) [6], 传毒媒介昆虫也是B型烟粉虱 (Bemisia tabaci) [6].Eu MV侵染紫果西番莲后, 起初叶片上有零星斑点, 继而发展为叶片出现坏死斑, 叶片和花朵畸形[34].烟粉虱是Eu MV的传毒媒介, 可在菜豆之间传毒, 但不能从西番莲传毒到菜豆或其它西番莲[34].Eu LCV和Pa LCu Gd V主要侵染紫果西番莲, 引起花叶和叶片畸形, 其传播途径不明[10].
1.3 香石竹潜隐病毒属
西番莲潜隐病毒 (passion fruitlatent virus, PLV) 是乙型线形病毒科 (Betaflexiviridae) 香石竹潜隐病毒属 (Carlavirus) 成员[35].PLV广泛侵染细柱西番莲 (Passiflora suberosa) 、蓝冠西番莲 (Passiflora caerulea) [36]、粉花西番莲 (Passiflora incarnata) 与拳卷西番莲 (Passiflora cincinnata) 的杂交后代[37-38]、紫果西番莲 (Passiflora edulis) 以及白花西番莲 (Passiflora subpeltata) [39].病毒粒子为弯曲的线条状, 无包膜, 长度600~700 nm, 直径12~13 nm, 基因组为单分体, 包含一条线形单链正义RNA.
不同类型的西番莲被PLV侵染后, 均表现为不明显的系统性花叶症状, 气温较低时, 老叶呈斑驳状。PLV通过蚜虫以非持久性方式传播, 具体蚜虫类型有待证实[6].
1.4 黄瓜花叶病毒属
黄瓜花叶病毒属 (Cucumovirus) 是雀麦花叶病毒科 (Bromoviridae) 成员, 是一种三分体单链RNA病毒, 包括4个RNA片段, 分别编码不同的蛋白, 病毒粒子为等轴对称的二十面体[40-41].在有些黄瓜花叶病毒 (Cucumber mosaic virus, CMV) 中, 除含有病毒本身的4段RNA外, 还含有一种分子量更小的卫星RNA.卫星RNA不能独立侵染和复制, 完全依赖于CMV的辅助作用, 但卫星RNA能干扰CMV复制, 降低CMV浓度[40,42].CMV寄主广泛, 能够侵染1 200多种植物, 已经报道的CMV株系或分离物有100个以上[41,43].中国从西番莲中分离的CMV株系有5种 (PE、PE2、PC、PF和PEf) [6,44], 株系PE、PE2、PC和PF属于CMV亚组Ⅰ;株系PEf属于CMV亚组Ⅱ。亚组Ⅰ在田间侵染的西番莲中占绝对优势[6].
紫果西番莲被CMV侵染后, 叶片呈典型花叶。在嫩叶上有明显黄点或黄斑, 顶部新抽出嫩叶发生卷曲;老叶有褪绿及黄化现象;病株矮化, 生长不良, 花、果明显减少。黄果西番莲被CMV侵染后也呈典型花叶, 老叶上有黄化斑点, 但病株无明显矮化现象[6,45].此外, 上述2种西番莲类型的果实感染CMV后, 均有石果症状[45].自然条件下, CMV主要通过桃蚜 (M.persicae) 、棉蚜 (A.gossypii) 以非持久性方式传播[6,45].
1.5 芜菁黄花叶病毒属
西番莲黄花叶病毒 (Passion fruit yellow mosaic virus, Pa YMV) 是芜菁黄花叶病毒科 (Tymoviridae) 芜菁黄花叶病毒属 (Tymovirus) 成员, 目前只在巴西和哥伦比亚有相关报道[46-47], 从巴西分离的Pa YMV株系只侵染西番莲科植物[46], 而哥伦比亚株系除侵染西番莲外, 还能侵染3种酸浆属植物[47].该病毒粒子为等轴对称的二十面体, 无包膜, 直径约30 nm, 病毒核酸为单分子线形正义单链RNA[6].
Pa YMV侵染西番莲引起叶片皱褶、亮黄色的网状花叶。Pa YMV巴西株系可通过南美叶甲 (Diabrotica speciosa) 传播, 传毒率较低, 而哥伦比亚株系则不能通过南美叶甲传播[6].
1.6 柑橘粗糙病毒属
西番莲绿斑病毒 (Passion fruit green spot virus, PFGSV) 是芜菁黄花叶病毒科 (Tymoviridae) 柑橘粗糙病毒属 (Cilevirus) 成员, 因成熟果实上有2~5 mm大小的绿色斑点而得名[48].该病毒粒子呈短棒状, 有包膜, 长度100~200 nm, 直径50~70 nm[49].
西番莲被PFGSV侵染后, 老叶沿着叶脉出现下陷的绿色坏死病斑, 病害严重时, 病斑逐渐覆盖、环绕枝蔓, 导致植株死亡, 甚至毁园[6].PFGSV通过紫红短须螨 (Brevipalpus phoenecis) 传播, 病毒一般只存在于紫红短须螨咬食区域, 而不会系统性转移至植株的其它部位[50].
1.7 烟草花叶病毒属
西番莲花叶病毒 (Maracuja mosaic virus, MarMV) 是帚状病毒科 (Virgaviridae) 烟草花叶病毒属 (Tobamovirus) 成员。目前报道的Mar MV有多个分离物, 分别在印度[51]、秘鲁[52]、美国[53-54]分离得到。秘鲁分离物Mar MV-P早已被确定是烟草花叶病毒属成员之一[52], 随后, 通过基于病毒基因组的系统发育分析证实美国毒系Mar MV-FL和Paf MV与MarMV-P一样, 都属于烟草花叶病毒[54].该病毒粒子呈杆状, 长度约320 nm, 直径约18 nm[6].
Mar MV侵染西番莲后引起花叶、叶片皱缩。该病毒并不通过传毒媒介传播, 而是通过植株之间的接触以及农业生产措施造成的机械损伤进行传播[6].
1.8 线虫传多面体病毒属
番茄环斑病毒 (Tomato ringspot virus, To RSV) 是豇豆花叶病毒科 (Comovirinae) 线虫传多面体病毒属 (Nepovirus) 成员, 病毒粒子是等轴颗粒, 直径为25~30 nm[6].To RSV侵染紫果西番莲仅在秘鲁有过报道, 而且是和Mar MV复合侵染, 单独侵染引起的病症尚不清楚, 主要通过美洲剑线虫 (Xiphinema americanum) 传播[55].
1.9 其它侵染西番莲的病毒
1.9.1 西番莲脉明病毒 (Passion fruit vein clearing virus, Pa VCV)
Pa VCV是一种弹状病毒 (Rhabdoviridae) , 长约300 nm, 直径约70 nm, 能侵染紫果西番莲、黄果西番莲 (Passiflora edulis f.flavicarpa) 和苹果西番莲 (Passiflora maliformis) [6].
该病毒的寄主范围和携毒载体 (昆虫) 尚不明确, 侵染西番莲除导致叶片明脉外, 还会引起叶面积和果实体积减小[56], 主要通过嫁接传播[57].
1.9.2 紫果西番莲花叶病毒 (Purple granadilla mo-saic virus, PGMV)
PGMV只侵染紫果西番莲, 病毒形态为等轴颗粒状, 直径约24 nm, 还未明确分类[6,58].PGMV侵染引起紫果西番莲轻度线状花叶, 果实变小、畸形、木质化[6,58], 气温较低的季节症状较为明显, 夏季症状几乎消失。该病毒可通过南美叶甲 (Diabrotica speciosa) 传播[6].
2、西番莲病毒检测技术
病毒种类的准确检测和鉴定是开展西番莲病毒病防治相关研究的前提条件, 已报道的检测方法主要有生物学检测、电子显微镜检测、血清学检测和分子生物学检测等技术。
2.1 生物学检测
生物学检测法又叫指示植物检测法, 其原理是将待测病毒接种到对该病毒敏感、短时间内即可表现出典型症状的寄主植物, 依据观察到的典型症状来判定病毒种类。该方法成功应用于黄瓜花叶病毒[8]、乌干达西番莲病毒[26]等病毒的鉴定, 主要指示植物有烟草 (Nicotiana tabacum L.cv.Xanthi, N.benthamiana Domin, N.glutinosa L.) 、豇豆 (Vigna unguiculata L.Walp) 、藜 (Chenopodium quinoa Willd.) 、黄瓜 (Cucumis sativus L.) 和西葫芦 (Cucurbitapepo L.) [59].该法易受季节和环境因素影响, 检测周期较长, 且容易出现假阳性[60].
2.2 电子显微镜检测
电子显微镜技术广泛应用于乌干达西番莲病毒[26]、大戟花叶病毒[34]和西番莲黄花叶病毒[47]检测, 是判断病毒是否存在的最直观的检测方法。该法不仅可以鉴别病毒类型, 还能观察到形态特征、特异结构等, 其检测结果是否准确取决于电镜的质量和标本制作效果;此外, 检测人员必须具备成熟的电镜操作经验和扎实的病毒学知识[60].
2.3 血清学检测
血清学检测是应用抗原抗体在体外特异性结合待测植物病毒, 产生血清学反应, 以此来鉴定病毒的方法。主要包括酶联免疫吸附法、免疫胶体金技术和快速免疫滤纸法, 其中以酶联免疫吸附法应用最为广泛, 该法灵敏度高、操作简便、检测周期短、结果准确, 但特异性抗体制作过程复杂, 而商用抗体价格较高[60].目前, 针对西番莲病毒豇豆蚜传花叶病毒[16]、乌干达西番莲病毒[26]、大戟花叶病毒[34]和黄瓜花叶病毒[44]等有可用的血清学检测试剂或抗体。
2.4 分子生物学检测
分子生物学检测是通过检测病毒核酸来判断病毒存在与否, 最常用的检测方法是聚合酶链反应 (Polymerase Chain Reaction, PCR) 和逆转录–聚合酶链反应 (Reverse Transcription-PCR, RT-PCR) , 其中RT-PCR应用于豇豆蚜传花叶病毒、西番莲斑驳病毒、西番莲Y病毒和黄瓜花叶病毒等多种病毒的检测[16,21].PCR、RT-PCR灵敏度高、特异性强, 能检测到微量样品中的病毒, 还能用于类病毒或不稳定病毒的检测[60].
3、西番莲病毒病防治
迄今为止, 国内外已报道了26种侵染西番莲的植物病毒。大部分病毒引起的西番莲病害只是在少数国家有零星发生, 而豇豆蚜传花叶病毒引起的西番莲木质化病毒病在尼日利亚、中国、澳大利亚、巴西和乌干达等国家均有发生, 且对生产造成严重危害, 是全球西番莲产业发展的主要限制因素之一, 豇豆蚜传花叶病毒防治相关研究也最为系统[1].下面以豇豆蚜传花叶病毒病为例, 介绍西番莲病毒病的防治方法。
3.1 选育抗病品种
选育并推广抗病品种是防治园艺植物病害最经济有效的措施, 在豇豆蚜传花叶病毒病防治方面也得到了应用。
3.1.1 杂交育种
Freitas等[61]以抗木质化病毒 (豇豆蚜传花叶病毒) 的野生西番莲资源setacea为父本, 商业化品种紫果西番莲为母本, 先得到种间杂种, 然后从回交4次的重组同类系中筛选得到了抗豇豆蚜传花叶病毒病的株系。Santos等[62]以setacea和紫果西番莲作为杂交组合, 同样获得了抗性杂种;随后, Freitas等[63]的研究表明, 以杂交或者回交后代为父本, 与紫果西番莲再次回交后得到的抗性株系, 其果实品质和产量更优。
3.1.2 实生选种
Cerqueira-Silva等[64]对黄果西番莲72个商业化品种的实生苗株系进行了田间抗病性鉴定, 通过分析各株系在人工接种豇豆蚜传花叶病毒后的病情指数和单株产量, 最终筛选获得了UESB-A1、UESB-B2、UESB-A21和UESB-E31这4个抗性较好的种质。
3.1.3 分子育种
通过病毒外壳蛋白基因遗传转化手段获得病毒抗性的方法已被广泛应用, 将改法应用于豇豆蚜传花叶病毒病育种方面也取得了成功。Trevisan等[65-66]将豇豆蚜传花叶病毒外壳蛋白基因CP导入黄果西番莲, 在黄果西番莲分别接种3种不同的豇豆蚜传花叶病毒株系, 结果表明转基因植株对这3个病毒株系都具有抗性;Correa等[67]构建CP基因的植物表达载体, 并通过转化翅茎西番莲, 获得了21个转基因株系, 经过4次接种鉴定, 有2个株系完全没有病症, 其它株系也有不同程度的抗性。
3.1.4 体细胞杂交
体细胞融合可以克服远缘杂交不亲和的障碍, 实现抗性基因在亲缘关系较远的物种之间转移[68].从细柱西番莲[69]、BGP029和BGP 168[70]等野生抗性资源的叶肉组织中分离原生质体, 与商业化主栽品种进行体细胞杂交, 有望获得抗性品种。
3.2 农艺防治
生产实践证明, 一些栽培技术措施可以有效减轻木质化病毒造成的危害[1,71], 主要包括: (1) 脱毒苗木的繁育与推广; (2) 对于新建果园, 仔细观察新植苗木5个月后的生长情况, 发现疑似被病毒侵染的植株, 及时清除; (3) 修剪时注意消毒工具, 避免病毒通过机械传播; (4) 避免在果园附近种植豆科植物, 减少病毒传染源。
3.3 化学防治
蚜虫是豇豆蚜传花叶病毒的传毒昆虫, Fischer等[6]曾尝试采用化学农药防治蚜虫来预防该病害, 由于蚜虫以非持久性方式传毒, 啃食叶片几秒钟就能传播病毒, 因此这一方法收效甚微。苯并噻二唑 (acibenzolar-S-methyl) 是一种植物诱导抗病药剂, 防治豇豆蚜传花叶病毒病效果较好, 施用浓度为0.025和0.05 g/L时, 接种病毒的植株表现症状的叶片数目分别减少至对照的17%和22%.该药剂的作用机理可能是通过提高植物叶片中几丁质酶和β-1, 3葡聚糖酶的活性来提高植物对病毒的抗性[72].
4、问题与展望
国内西番莲病毒病最早发生在台湾地区, 随后在福建、广东和海南等西番莲产区也有报道, 主要病原为黄瓜花叶病毒。20世纪80年代以来, 病毒病对各地西番莲产业都造成了一定程度的危害, 但国内病毒病相关研究较少。
Chang[9]对西番莲斑驳病毒进行了鉴定, 并分析了该病毒与其它马铃薯Y病毒的遗传差异。徐平东等[43-44]、郑冠标等[45]对侵染西番莲的黄瓜花叶病病毒进行了鉴定和分型。王海河等[73]克隆了黄瓜花叶病毒西番莲致死分离物 (CMV-PE) RNA3的c DNA全长, 发现使西番莲症状相似的CMV分离物的非编码区具有很高的序列同源性, 非编码区序列与症状具有相关性。刘志昕等[74]尝试通过转基因手段导入黄瓜花叶病毒外壳蛋白基因来获得抗病植株, 但未见后续研究报道。总体而言, 国内西番莲病毒病相关研究多停留在病毒鉴定、分类层面, 而病毒致病机理、抗病育种等相关研究尚未取得突破性进展。
近年来, 贵州、广西、云南等省份将西番莲种植作为“短、平、快”的精准扶贫项目进行推广, 西番莲产业迎来了新的发展机遇。病毒病的防治对于西番莲产业的健康发展有着重要意义, 产业健康发展需要通过以下研究来加以保障: (1) 研发高效脱毒技术, 推广无病毒苗木, 提高病害预防意识; (2) 全面调查病毒病发生情况, 掌握病害流行规律, 为农艺防治、化学防治提供参考; (3) 应用现代分子生物学技术鉴定病毒, 了解其致病机理, 为抗病分子育种提供理论依据; (4) 加强抗病资源筛选和抗病种质创制相关工作, 为抗病品种选育奠定基础。通过开展以上研究, 可最大程度地减轻病毒病危害, 提高果农和种植企业的经济收益。
参考文献
[1]Cbm C S, Ldhcs C, Souza A P, et al.A history of passion fruit woodiness disease with emphasis on the current situation in Brazil and prospects for Brazilian passion fruit cultivation[J].European Journal of Plant Pathology, 2014, 139 (2) :261-270.
[2]Ocampo J, Arias J C, Urrea R.Interspecific hybridization between cultivated and wild species of genus Passiflora L.[J].Euphytica, 2016, 209 (2) :395-408.
[3]李莉萍。西番莲综合开发利用研究进展[J].安徽农业科学, 2012, 40 (28) :13840-13843, 13846.
[4]吴均秀。浅谈西番莲及其发展前景[J].广西热作科技, 1994, 50:43-46.
[5]王秀荣, 段安安, 许玉兰。国内西番莲引种栽培现状及改良思路[J].西南林学院学报, 2003, 23 (2) :88-91.
[6]Fischer I H, Rezende J A M.Diseases of Passion flower (Passiflora spp.) [J].Pest Technology, 2008, 2 (1) :1-19.
[7]陈光禄。台湾西番莲生产现状及其栽培管理[J].东南园艺, 1991 (4) :26-27.
[8]徐平东, 李梅。福建西番莲病毒病的发生及其病原黄瓜花叶病毒亚组鉴定[J].植物保护学报, 1999, 26 (1) :50-54.
[9]Chang C A.Characterizarion and comparation of passion fruit mottle virus, a newly recognized potyvirus, with passion fruit woodiness virus[J].Phytopathology, 1992, 82:1 358-1 363.
[10]Cheng Y H, Deng T C, Chen C C.First report of Euphorbia leaf curl virus and Papaya leaf curl Guangdong virus on Passion Fruit in Taiwan[J].Plant Disease, 2014, 98 (12) :1746.
[11]谢丽雪, 张小艳, 郑姗, 等。侵染西番莲的夜来香花叶病毒的分子鉴定及特异性检测[J].中国农业科学, 2017, 50 (24) :4 725-4 734.
[12]Jawaid A, Khan A J, Dijkstra J.Plant viruses as molecular pathogens[M].USA, Food Products Press (The Haworth Press Inc.) , NY, 2002:530.
[13]Mc Knight T.The woodiness disease of the passion vine[J].Queensland Journal of Agricultural Society, 1953, 10:4-35.
[14]Martini C K H.Some properties of the virus causing'woodiness'of passion fruit in Western Nigeria[J].Annals of Applied Biology, 1962, 50:163-158.
[15]Iwai H, Ohmori T, Kurokawa Y, et al.New report of passion fruit woodiness virus in Japan[J].Annals of the Phytopathological Society of Japan, 1996, 62:459-465.
[16]Wylie S J, Jones M G.The complete genome sequence of a Passion fruit woodiness virus isolate from Australia determined using deep sequencing, and its relationship to other potyviruses[J].Archives of Virology, 2011, 156 (3) :479-482.
[17]Fukumoto T, Nakamura M, Wylie S J, et al.Complete nucleotide sequence of a new isolate of passion fruit woodiness virusfrom Western Australia[J].Archives of virology, 2013, 158 (8) :18-21.
[18]Nascimento A V S, Santana E N, Braz A S K, et al.Cowpea aphid-borne mosaic virus (CABMV) is widespread in passion fruit in Brazil and causes passion fruit woodiness disease[J].Archives of Virology, 2006, 151 (9) :1 797-1 809.
[19]González-segnana L R, Ramirez d L M, Mello A P O A, et al.First report of Cowpea aphid-borne mosaic virus on sesame in Paraguay[J].Plant Disease, 2011, 95 (5) :613.
[20]de Wijs J J.A virus causing ringspot of Passiflora edulis in the Ivory Coast[J].Annals of Applied Biology, 1974, 77 (1) :33-40.
[21]Parrella G, Castellano M A.Passiflora chlorotic spot a disease caused by a strain of Bean yellow mosaic virus in Passilfora caerulea in Italy[J].Journal of Plant Pathology, 2002, 84 (2) :139.
[22]Parrella G, Lanave C.Identification of a new pathotype of Bean yellow mosaic virus (BYMV) infecting blue passion flower and some evolutionary characteristics of BYMV[J].Archives of Virology, 2009, 154 (10) :1 689-1 694.
[23]Benscher D, Pappu S S, Niblett C L, et al.A strain of soybean mosaic virus infecting Passiflora spp.in Colombia[J].Plant Disease, 1996, 80 (3) :258-262.
[24]Parry J N, Davis R I, Thomas J E.Passiflora virus Y, a novel virus infecting Passiflora spp.in Australia and the Indonesian province of Papua[J].Australasian Plant Pathology, 2004, 33 (3) :423-427.
[25]Iwai H, Yamashita Y, Nishi N, et al.The potyvirus associated with the dappled fruit of Passiflora edulis in Kagoshima prefecture, Japan is the third strain of the proposed new species East Asian Passiflora virus (EAPV) phylogenetically distinguished from strains of passion fruit woodiness virus[J].Archives of Virology, 2006, 151 (4) :811-818.
[26]Ochwo-Ssemakula M, Sengooba T, Hakiza J J, et al.Characterization and distribution of a Potyvirus associated with passion fruit woodiness disease in Ugan da[J].Plant Disease, 2012, 96 (5) :659-665.
[27]Abdullah N, Ismail I, Pillai V, et al.Nucleotide Sequence of the Coat Protein Gene of the Malaysian Passiflora Virus and its 3'Non-Coding Region[J].American Journal of Applied Sciences, 2009, 6 (9) :1 633-1 636.
[28]Chiemsombat P, Prammanee S, Pipattanawong N.Occurrence of Telosma mosaic virus causing passion fruit severe mosaic disease in Thailand and immunostrip test for rapid virus detection[J].Crop Protection, 2014, 63 (5) :41-47.
[29]Watanabe S, Ruschel R, Marrero G, et al.A distinct lineage of Watermelon mosaic virus naturally infects honohono orchid (Dendrobium anosmum) and Passion fruit (Passiflora edulis) in Hawaii[J].New Disease Reports, 2016, 34:13.
[30]Brown J K, Bird J, Fletcher D C.First report of passiflora leaf mottle disease caused by a whitefly-transmitted geminivirus in Puerto Rico[J].Plant Disease, 1993, 77 (12) :1 264.
[31]Brown J K, Bird J.Introduction of an exotic whitefly (Bemisia) vector facilitates secondary spread of Jatropha mosaic virus, a geminivirus previously vectored exclusively by the Jatropha biotype[M].UK, Intercept Publications, Wimborne, 1996:351-353.
[32]Novaes Q S, Astua J F, Yuki V A, et al.Partial characterization of a bipartite begomovirus infecting yellow passion flower in Brazil[J].Plant Pathology, 2003, 52 (5) :648-654.
[33]Vaca-Vaca J C, Carrasco-Lozano E C, López-López K.Molecular identification of a new begomovirus infecting yellow passion fruit (Passi flora edulis) in Colombia[J].Archives of Virology, 2017, 162 (2) :573-576.
[34]Polston J E, Londono M A, Cohen A L, et al.Genome Sequence of Euphorbia mosaic virus from Passion fruit and Euphorbia heterophylla in Florida[J].Genome Announcements, 2017, 5 (9) :e01 714-e01 716.
[35]Spiegel S, Zeidan M, Sobolev I, et al.The complete nucleotide sequence of Passiflora latent virus and its phylogenetic relationship to other carlavirus[J].Archives of Virology, 2007, 152:181-189.
[36]Brandes J, Wetter C.Studies on the charac teristics and relationships of Passiflora latent virus[J].Phytopatologiese Zeitschrift, 1963, 49:61-70.
[37]St Hill A A, Zettler F W, Elliot M S, et al.Presence of passiflora latent virus and a serologically distinct strain of Maracuja mosaic virus in Passiflora spp.in Florida[J].Plant Disease, 1992, 76 (8) :843-847.
[38]Hicks R G T, Mohamed M E, Blakesley D.Passiflora latent carlavirus in European collections of ornamental Passiflora[J].Journal of Phytopathology, 1996, 144 (4) :203-205.
[39]Pares R D, Gunn L V, Keskula E N, et al.Occurrence of Passiflora latent carlavirus in cultivated and wilt Passiflora species in Australia[J].Plant Disease, 1997, 81:248-350.
[40]邱艳红, 王超楠, 朱水芳。黄瓜花叶病毒致病性研究进展[J].生物技术通报, 2017, 33 (9) :10-16.
[41]Palukaitis P, Roossinck M J, Dietzgen R G, et al.Cucumber mosaic virus[J].Advances in Virus Research, 1992, 41:281-348.
[42]Palukaitis P, Garcia-Arenal F.Cucumoviruses[J].Advances in Virus Research, 2003, 62:241-323.
[43]徐平东, 谢联辉。黄瓜花叶病毒亚组研究进展[J].福建农业大学学报 (自然科学版) , 1998, 27 (1) :82-91.
[44]徐平东, 李梅, 林奇英, 等。侵染西番莲属 (Passiflora) 植物的五个黄瓜花叶病毒分离物的特性比较[J].中国病毒学, 1999, 14 (1) :73-79.
[45]郑冠标, 高乔婉, 张曙光, 等。鸡旦果花叶病病原病毒的鉴定[J].华南农业大学学报, 1987, 8 (2) :40-44.
[46]Crestani, O A, Kitajima, E W, Lin M T, et al.Passion fruit yellow mosaic virus, a new Tymovirus found in Brazil[J].Phytopathology, 1986, 76 (9) :951-955.
[47]Morales F J, Lozano I, Castano M, et al.Partial characterization of a Tymovirus infecting passion fruit in Colombia, South America[J].Journal of Phytopathology, 2002, 150 (4-5) :292-296.
[48]Locali-Fabris E C, Freitas-Astua J, Souza A A, et al.Complete nucleotide sequence, genomic organization and phylogenetic analysis of Citrus leprosis virus cytoplasmic type[J].Journal of General Virology, 2006, 87 (9) :2721-2729.
[49]Kitajima E W, Rezende J A M, Rodrigues J C V, et al.Green spot of passion fruit, a possible viral disease associated with infestation by the mite Brevipalpus phoenicis[J].Fitopatologia Brasileira, 1997, 22:555-559.
[50]Kitajima E W, Rezende J A M, Rodrigues J C V.Passion fruit green spot virus vectored by Bevipalpus phoenecis (Acari:Tenuipalpidae) on passion fruit in Brazil[J].Experimental and Applied Acarology, 2003, 30:225-231.
[51]Mali V R, Vyanjane N T.Occurrence of tobacco mosaic virus on passionflower (Passiflora caerulea) [J].Indian Journal Mycology and Plant Pathology, 1980, 11:112-114.
[52]Fribourg C E, Koenig R, Lesemann D E.A new tobamovirus from Passiflora edulis in Peru[J].Phytopathology, 1987, 77 (3) :486-491.
[53]Song Y S, Min B E, Hong J S, et al.Molecular evidence supporting the confirmation of Maracuja mosaic virus as a species of the genus Tobamovirus and production of an infectious c DNA transcript[J].Archives of Virology, 2006, 151 (12) :2337-2348.
[54]Song Y S, Ryu K H.The complete genome sequence and genome structure of passion fruit mosaic virus[J].Archives of Virology, 2011, 156 (6) :1 093-1 095.
[55]Koenig R, Fribourg C E.Natural occurrence of Tomato ringspot virus in Passiflora edulis in Peru[J].Plant Disease, 1986, 70 (3) :244-245.
[56]Kitajima E W, Chagas C M, Crestani O A.Virus and mycoplasma-associated diseases of passion fruit in Brazil[J].Fitopatol Brasileira, 1986, 11:409-432.
[57]Pares R D, Martin A B, Morrison W.Rhabdovirus-like particles in Passion fruit[J].Australasian Plant Pathology, 1983, 12 (3) :51-52.
[58]Oliveira C R B, Marinho V L A, Astolfi Fo S, et al.Purification, serology and some properties of the purple granadilla (Passiflora edulis) mosaic virus[J].Fitopatologia Brasileira, 1994, 19 (3) :455-462.
[59]Parrella G, Sorrentino D.Identification of a cucumber mosaic virus isolate from Passiflora edulis in Southern Italy and validation of subgroup identification by In silico restriction fragment length polymorphism[J].Journal of Phytopathology, 2009, 157:762-767.
[60]陶源, 吴兴泉。植物病毒检测方法的研究进展[J].分子植物育种, 2017, 15 (7) :2 901-2 906.
[61]Freitas J C O, Viana A P, Santos E A, et al.Genetic basis of the resistance of a passion fruit segregant population to Cowpea aphid-borne mosaic virus (CABMV) [J].Tropical Plant Pathology, 2015, 40 (5) :1-7.
[62]Santos E A, Viana A P, Freitas J C O, et al.Resistance to Cowpea aphid-borne mosaic virus in species and hybrids of Passiflora:advances for the control of the passion fruit woodiness disease in Brazil[J].European Journal of Plant Pathology, 2015, 143:85-98.
[63]Freitas J C O, Viana A P, Santos E A, et al.Sour passion fruit breeding:Strategy applied to individual selection in segregating population of Passiflora resistant to Cowpea aphid-bornmosaic virus (CABMV) [J].Scientia Horticulturae, 2016, 211:241-247.
[64]Cerqueira-Silva C B M, Moreira C N, Figueira A R, et al.Detection of a resistance gradient to passion fruit woodiness virus and selection of'yellow'passion fruit plants under field conditions[J].Genetics and Molecular Research, 2008, 7 (4) :1 209-1 216.
[65]Trevisan F, Mendes B M J, Maciel S C, et al.Resistance to passion fruit woodiness virus in transgenic passion flower expressing the virus coat protein gene[J].Plant Disease, 2006, 90 (8) :1026-1030.
[66]Trevisan F, Mendes B M J, Maciel S C, et al.Resistance to passion fruit woodiness virus in transgenic plants of the yellow passion fruit expressing the viral coat protein gene[J].Acta Horticulturae, 2007, 738 (738) :495-499.
[67]Correa M F, Pinto A P C, Rezende J A M, et al.Genetic transformation of sweet passion fruit (Passiflora alata) and reactions of the transgenic plants to Cowpea aphid borne mosaic virus[J].European Journal of Plant Pathology, 2015, 143 (4) :813-821.
[68]Liu J H, Xu X Y, Deng X X.Intergeneric somatic hibridization and its application to crop genetic improvement[J].Plant Cell, Tissue and Organ Culture, 2005, 82 (1) :19-44.
[69]Maciel S D C, Nakano D H, Rezende J A M, et al.Screening of Passiflora species for reaction to Cowpea aphid-borne mosaic virus reveals an immune wild species[J].Scientia Agricola, 2009, 66 (3) :414-418.
[70]Cerqueira-Silva C B M, Jesus, O N, Oliveira E J, et al.Characterization and selection of passion fruit (yellow and purple) accessions based on molecular markers and disease reactions for use in breeding programs[J].Euphytica, 2015, 202 (3) :345-359.
[71]Prammanee S, Thumjamras S, Chiemsombat P, et al.Efficient shoot regeneration from direct apical meristem tissue to produce virus-free purple passion fruit plants[J].Crop Protection, 2011, 30 (11) :1 425-1 429.
[72]Parkinson L E, Crew K.S, Thomas J E, et al.Efficacy of acibenzolar-S-methyl (Bion) treatment of Australian commercial passion fruit, Passiflora edulis f.sp.flavicarpa, on resistance to Passion fruit woodiness virus (PWV) and activities of chitinase andβ-1, 3-glucanase[J].Australasian Plant Pathology, 2015, 44:311-318.
[73]王海河, 谢联辉, 林奇英。黄瓜花叶病毒西番莲分离物RNA3的c DNA全长克隆和序列分析[J].福建农业大学学报, 2001, 30 (2) :191-198.
[74]刘志昕, 魏源文。CMV-Pf-CP基因表达载体的构建及转化西番莲 (Passiflora edulis) 的研究[J].西南农业学报, 2001, 14 (4) :10-13.○R