������ʵ�����ӣ������ԣ��ȣ���������������ָͬһֲ��IJ�ͬ��λ������ͬ��̬����Ϊ��ʵ�����ӣ����������ڹ�ʵ�����Կ��Ե���ֲ����������ɢ���������ȷ��ȷ��������ͬ��̬����ѧ���ԣ�����ڽ����ܼ���Լ������ͬ��������ЧӦ���ԣ���ֲ����ʱ�������Ի����²�ȡ���շֵ�����ͷ��ע��Ӧ�Բߵ���Ҫԭ�����������ӵģ�����������Ҫ�ֲ��ڸɺ���ɺ�������Į����������������ǿ�ҵĵ�������ʵ�����Զ�ֲ����Ӧ�ɺ����ӻ��������������塣
����Ŀǰ�������������ѳ�Ϊ�������о����ȵ㣬�Ѿ����������ӵ��ȷ����������ԡ��������ӵ���̬�ṹ�������������ӵĴ����ص㡢�������Ӻ�������ֲ�������������Ӧ���ֵȷ���ȡ�ô����ɹ������������������γɵ���̬ѧ�����ȷ����о����٣������������������������Ӧ�Ĺؼ���
�����ϳ���ë�ˣ��ӣ�����ᡡ�������ã������ͣ����Ϊ��ë�����������ݱ�ֲ��ֲ������ǡ��������������ҹ����ֲ����½�������أ����к�ǿ�Ŀ��ɺ��Ϳ��μ��������ǻ�Įֲ���Ľ�Ⱥ�ֻ������֣��������ķ����ɳֲ��ϳ���ë���Ǿ��й�ʵ�����ԵĴ�����ֲ������У����¡��������ӣ������ȷ�����Ҳ��һ�������ڲ�ͬ�������Ӵ��ڹ����С�IJ�ͬ�����Ի������ͬ����ɢ��ֺ��ȷ���Ϊ������ͨ��Ұ����鷢�֣��ϳ���ë�˾��д����ġ��������ȷ����ص㣬��ֻҪ�������˾Ϳ����ȷ����Ӷ��γ��ˣ�����Ⱥֲ���С��һ��������Ӧ��ˮ�����Բ������������ԣ�����������������γɵ���̬ѧ�ں����б�Ҫ�о���
���������ϳ���ë��Ұ�������仯�������ǽ�ˮ����Ϣ������ã�������ֲ���С���ȷ�ʱ�������������Ϊ���ݣ����öԱȵķ������о����½������ϳ���ë����Ҫ�ֲ�����ͬ������Ⱥ������������������α仯�ģ���ͬ�������ͱ�������Ⱥ���Ƿ�һ�£��Ƿ���ֲ���С����������ϵ���Դ˷����������������γɵ�������̬ѧԭ���Էḻ��������������о���
�����������뷽��
�����������о����ſ�
�������������й��ڶ�����أ��½�������������ڽ��У�����ֲ���������ر�Ե��ǰɽ����Į�ش������ص�ɳĮ�ش�������°����′������°����′�Σ�����°����′������°����′�ţ��������������������������в�Ϊ��ԭ��ɳĮ����Ե����ɽ´�����ޣ�ɳĮԼռ�����������Σ����������������������������´������е��͵Ĵ�½�Ըɺ���ɺ��������������ļ�Ϊ�缾�����^��ʪ���ļ����ȣ�������������������ƽ������Ϊ�������棬�������¶ȣ����� �棬�����£��������� �棬���²���������棬���²�ƽ������ �����ϣ��¾���≥������·ݣ������У��������£�≥�����������£������������������棬��˪��Ϊ���������������䣬̫�����ܷ�����Լ��������ʣ��������������ʱ��Լ���������裬ƽ����ѩ���Ϊ��������������ۣ����ݡ�ɳĮ���꽵ˮ��Լ��������������������ȴ �� ������� ������ �ϣ��� �� ˮ �� �ģ����������������������꽵ˮ��һ�㣴������������������
�����������ֲ���Ժ���������ֲ��Ϊ�����������ǻ�Į���ص㡣ֲ������ϡ�٣�Ⱥ��ṹ���ơ��տơ�ʮ�ֻ��ơ����ƺͺ̱��Ƶ�ֲ��ϳ������ḻ��������Դ�����˶����Ļ�Įֲ��Ⱥ�����ͣ���Ϊ�����´���Į����Ϊ�����һƬ����
��������������ȡ������������
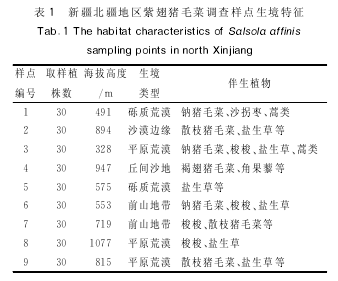
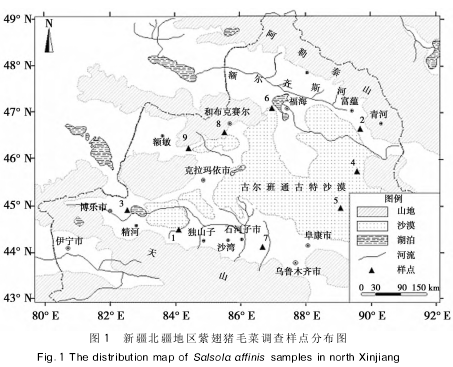
�������о������ϳ���ë�����½����������㷺�ֲ�����Ҫ���������ʻ�Į��ɳĮ��Ե�����ɳ�ء�ǰɽ�ش��ȵأ����Σ������������������������������о���������ر�Ե�����ع�ȡ�������㣬���������˸����ֵ���Ҫ�ֲ�����ͼ������
����������ȡ������
�������������꣱�������������ϳ���ë�����ӳ����ڣ���ͼ���еģ����о���ѡȡ�ϳ���ë�˵��ͷֲ����أ����Ʊ�ǡ��ⶨ��Ⱥ��ֲ��ڷ�����ߣ�����Ⱥ��ֲ�껮��Ϊ���С�С��������ͳ����Ŀ��Ȼ��ÿ��ֲ�꼶�����ȡ�������꣬�������ܹ�ȡ�������ꡣ��ֲ�����������ڳ�������װ���ܷ���д���ʵ���ҽ�һ��������
������������ʵ��̬�����������ⶨ
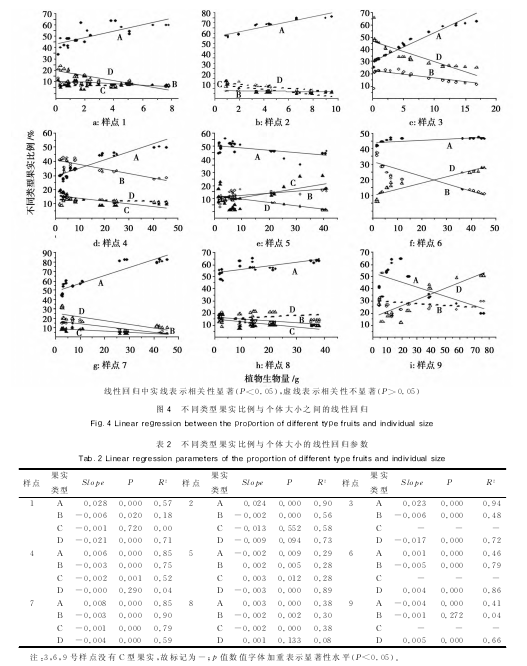
�������о��н��ϳ���ë�˴��л���Ƭ����ɢ�������ʵ��ÿ��������ѡȡ����С���ϳ���ë�˸��壳���꣬�۲��¼��ʵ�ϻ���Ƭ����̬����ķ����������ͬ����ʵ����״����ɫ������������ʽ�����α꿨�߷ֱ������ͬ����ʵ��������ʵ��ֱ���ͺ�ȣ��õ�ÿ����ʵ����ֵ����ʵ�����IJⶨ��ÿ������ʵ��������×�����ƽ��ֵΪ��ƽ�������ء��о��������㲻ͬ����ʵ��������̬��������һ�����죬���Խ����������ڹ�ʵ���ͷ�Ϊ�������ࣨͼ������ͳ������ֲ���ϲ�ͬ����ʵ���������ó���ͬ����ʵ��ռ�ı�����
����
����������ֲ���������IJⶨ
����ֲ�����ʵ���Һ�ÿ��ֲ��ֳ�Ӫ�����٣���������Ҷ������ֳ���٣���ʵ��������װ��ֽ���У��ڣ����������º�ɣ����裬Ȼ���ã�������������������ƽ��������ء�
�������������ݴ���
�������ãӣУӣӣ�����������������ʵ��ֱ���������ء�������ֲ�����������е����ط���������ϣ�壭����������Σϣ֣�������������С�����Բ��췽�����̣ӣģ����ж��رȽϣ�ȷ����ͬ������������IJ�������ˮƽ���У��������������Բ�ͬ��ʵ���ͱ�����ֲ������С֮��Ĺ�ϵ�������Իع��������ͼ����ͼ���ãϣ���������������ϣ�����ɣ�㣮����
��������������
���������������ڹ�ʵ���ͻ�������
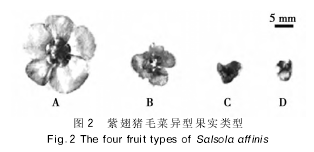
�������ݹ�ʵ��ɫ���ڲ�����������ʽ�����ᣨ�滨��Ƭ��ֱ������ʵ�����ص����������Բ����ԣ����ǽ��ϳ���ë�˵Ĺ�ʵ��Ϊ�����¡��á��ġ��������ͣ���������ֻ���ڣ����¡��ġ����ࡣ
��������ʵ�����Ϻ�ɫ��״�������ʵ�Ƚϱ������ں���ɫԲ�κ������ӣ�ö��
��������ʵ�����Ϻ�ɫ��״������ȣ�����С���ں���ɫԲ�κ������ӣ�ö��
��������ʵ���滨��Ƭ�ı��������Ϻ�ɫͻ�����״������ں���ɫ����Բ�����ӣ�ö��
��������ʵ���滨��Ƭ������״������ҹ�ʵ��ƽ����Բ�Σ��ں�����δ����Ļ�ɫֱ����Բ�����ӣ�ö��
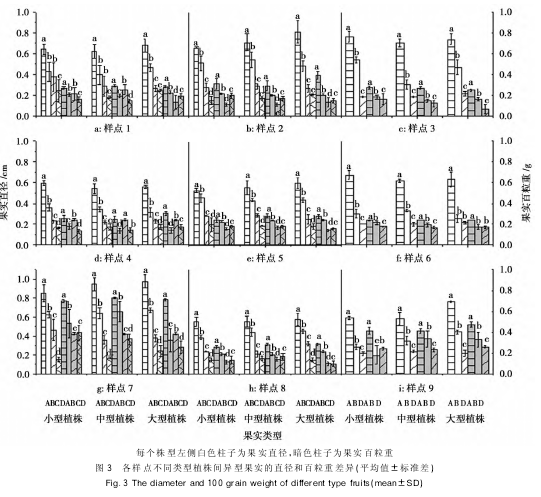
��������������ʵ��ֱ��������������ͼ����ʾ��ͨ��ͳ�ƣ���������ϳ���ë������ʵ����̬������������������֣��ڣ����������������������ȣ��������ڲ�ֲͬ�������ϵģ����ʵ��ֱ���������ţ��У��������������ʼ�С���ƣ�������Щ����Ĺ�ʵ�����ط��棬���㣲����������������������ʵ�����ز������ţ��У����������������������㣳�ʼ�С���ƣ����㣲���������ģ���ʵ�ȣ���ʵ�أ����㣷�Ĵ���ֲ���ϣ���ʵ�ȣ����أ����㣹��С��ֲ���Ϲ�ʵ�ĵ��������ڣ¡���ʣ�µģ��������У����㣶��С�͡�����ֲ���ϵģ����ʵ��ֱ���Ͱ����ض��������ţ��У��������������ҳ��½����ƣ�������ֲ��ģ��������ʵ�IJ��첻���ţ������㣱�ͣ��IJ�������ʵ�ڹ�ʵֱ�����������ϲ��첢�����š���Ȼ�ڲ�ͬ�����ڲ�ͬ�����ϴ���һ�����첻���ŵ����������������������������ڣ���������ʵ���۴�ֱ�����ǰ������϶��Dz������ŵģ��ʵݼ����ƣ�������ʵ��ֱ���Ͱ��������ǣ����ʵ����С�ġ�
������������ʵ���ͱ���������С֮��Ĺ�ϵ
����ͨ�������Բ�ͬ��Ⱥ�Աȷ������֣��ϳ���ë�����Ÿ�����С����������ʵ����������ʵ���������࣬����ʵ���͵ı�������ֲ���С��������ͬ�ı仯�������㣱���������������������У�����ʵ��ռ������ֲ���С�ʼ����ŵ�����أ��У�����������������ֲ���С�������������ʵ�����á�����ʵ����ֲ���С���³ʸ���ع�ϵ��
�������ڲ��������У����������IJ������ֲ�ͬ������ԣ����㣵�ͣ�����ֲ����������������ʵ�ı��������½����ƣ�����ʵ�����㣶�ͣ�������ֳ���ֲ���С�ʼ���������أ��У����������������Ͻ�������ϳ���ë�˹�ʵ�������������ֲ���С�����ԣ���������ԴͶ����ͬ����£���ͬ��������֮��Ҳ������ԴȨ�⡣�����ۺϽ��˵����ֲ�����Խ�ԣ���ʵ��Ͷ���Խ�ߣ����ԣ¡��á�����ʵ��Ͷ��ή�͡�
����������������
������Į�ɺ����꣬�¶ȡ�ʪ���ڣ����ڱ仯���ң����併ˮʱ������ʴռ�ֲ��������Ըߡ�����Ͽ���Ļ��������ࣱ����ֲ���ʵ�����ӣ����γɶ�������̬��������������Ӱ�촫�����ȷ���Ϊ����ߴ���ʺ���Ӧ�ԡ���Ŀǰ�о�������������ֲ�ﵥ���ͬʱͨ���������������ӣ����ڴ����ĺͲ��״����ģ������������ӷֱ����ȷ����Դ����Ų����ߺ�������������ڿռ��ϣ����ڴ�����������ɢ������ͬ�����ӣ���ɢ��ֲ����ĸ����Χ���ܶ��������ܼ���Լ��ͬ����������ʱ���ϣ������������̶߳ȵIJ��죬ʹ�ø������ڶ��Ļ�Į�����У���ʹ�������ݵ����˻���Ҳ��ȫ���ȷ������ߣ��Ӷ�������ڻ����������µ���Ⱥ�������֤���ֵ������о���Աͨ���۲⣱����Ȼ��Ⱥ���֣��ϳ���ë�˵����ͬʱ���������¡�����ʵ�����У�������ʵ���ڴ��������ӻ����ϸߣ�������ʵ��ɢ�����������������Ȼ�����о�ͨ��ȫ�����������صģ������㷢�֣��ϳ���ë�˵��겻����ͬʱ���������¡�����ʵ�����һ��ձ���ڽ��ڣ�������ʵ�е�С���ʵ����ʹ�ù�ʵ���ʹ�κ�ҵ��˵ģ������ӵ����ڵģ��࣬�ܹ�ͬʱ���������¡��á�����ʵ����Ⱥռ���۲�����ģ��������������������������Էdz��ձ飬�������Ż����仯��������̬�仯���ḻ��
�������ϳ���ë��ֲ���ϣ���ͬ����ʵ���ص�����������Ӧ������ѧ���ܡ��������ɵ��˵��о�������ԱȽ����⣴���ʵ�Ĵ������塣
��������ʵ��Ҫ�ֲ���ֲ����ϲ�����Ϊ���죬���״�ĸ�����룬��紫������Զ�ĵط�����Ϊ“������”ռ���������Σ������Լ������淶Χ������ʵ�ֲ���ֲ������ϲ����������״�ĸ�����룬��紫������Զ�ĵط�����Ϊ����ʵ��“����������”������ʵ��Ҫ�ֲ���ֲ������²���������״�ĸ�������룬����Ƭ��״������;�����ֻ��ɢ����ĸ��ײ����Ǹ�������Ϊ“������”��������Ⱥ��ԭ�����������ռ�ݣ�����ʵ����ֲ��ڹ�֦����˼�������ͨ�����ʷ����߲�δ���죬�һ���Ƭ��������Χ�γɽϺ�ʵ����֯�ṹ��������̬���������������������������ӿ��б���ϳ�ʱ�䣬�ھ����������졢�����������ô�������֮���п��ܾ߱��ȷ�����������Ϊֲ��������“������”���ڡ������о����֣������������ȷ��ʺ��������Բ������ԣ���˿����ƶ������������Ǹ�������Ӧ��Į����Ҫ�ֶΡ�
�������ڴ�������ʷ��Ȩ�⣬ֲ���Ӫ����������ֳ������������ٽ�����Լ��ϵ�����ֳ���ֳ����������������������ڷ�ֳ�����ڲ���ֳ����֮�䣬�����Ǿ��������Թ�ʵ�����ӣ��IJ�ͬ����֮������α仯�ļ��б�Ҫ���������
�����ϳ���ë�����ڣ�����ֲ����ã�Բߣ����������������ӣ������ܴ��ռ�зḻ������������Դ���ƣ��������Ͻ��ͻ��������Ա仯��в�ȶ���ȺΣ�����������������������������ԴͶ����仯���棬����ɵ��о�������ֲ������������������ռ���������࣬����������ռ�������٣�����������ռ����û�����ű仯�����о�ͨ�������Բ�ͬ��Ⱥ�Աȷ��������ϳ���ë�����Ÿ�����С����������ʵ���������������������࣬���ι�ʵ���͵ı�������ֲ���С��������ͬ�ı仯������ʵ��ֲ���С������ع�ϵ������ֲ���С��������������ǣ������ӣ����ǣá�����ʵ����ֲ���С���³ʸ���ع�ϵ����Ȼ������������������������ϳ���ë����������������ֲ���С�����ԣ���������ԴͶ����ͬ����£���ͬ��������֮��Ҳ������ԴȨ�⡣
�����ԣ��������ָ��ֲ��Դ�С��ͬ�����ӵ�ѡ���ǶԶ���������һ����Ӧ�����о������ϳ���ë�˴�ֲ������ܴ�����ģ���ʵ�����ͣ¡��á�����ʵ�����������ʵ���������ֿ��Դ�ʹ�ϳ���ë���ڻ����������˵�����£�Ѹ��ռ��ԭ�������ռ䣬����������ɢ���ڿռ����ӱܻ���������ֲ���Ӱ�죬�Ӷ��ڱ���ԭ����Ⱥ�Ļ����ϴٽ���Ⱥ�����������š���Сֲ������У¡��á�����ʵռ�кܴ�ı��أ���ЩСֲ��IJ�����Ҫ���ڽ�ˮ���ͣ�ֲ���������ޡ��������ڴﵽ�ϳ���ë�������ȷ��Ľ�ˮ����������ֲ��Ӫ������ʱ��������¡����ڣ���ʵ�ȷ������Ե��ڣ���ʵ�������ڻ��ȣ��á�����ʵ�ȷ��ʼ��ͣ��γ����ӿ⣬�����Ϊ�ڻ��������������Ͽ�����£��ϳ���ë��С��ֲ��ķ�ֳ���Ը�������“����”��ʵ����Ⱥ��������ƻ���Ļָ���������Ⱥ�������
�������⣬���Ǵ��ϳ���ë�����Ӵ�С��ֲ���С�����ϵ���Է��֣����Ӵ�С�仯�ܻ���в�ȺͲ�����ǿ��Ӱ�죬�¡��á��������Ӳ���������ֲ��Ӫ��״�����������ң��������ȷ��ʣ��������ڣ��á����������ȷ���С�ڣ����������������������������Ȼ�ӿ�����Щ�¡��á��������ӳ����ȷ����������Ⱥ������ά���������ף�����ҲҪ�����ڻ������������£�ֲ��������С�������˴�����Ч��������ʱ��Ч�����ӣ��Ƕ���Դ��һ���˷ѣ�����֮��������Щ�������������Ӿ���һ���ȷ�����������˸�ֲ��Բ���������Ӧ����ʵ��������ĸ��ЧӦ�����Ŵ���Ӱ�죬��Щ�������������Ӻ����������������ܷ�ָ�����ֲ�������������Ե����⼫�б�Ҫ�����о�����Ϊ����ȫ���˽�������ڻ�Į�Ͽỷ���㷺�ֲ��Ļ�����
�����ο����ף�
�����ۣ��ݣ֣�����塡�ġ��̣��ԣ�塡������������������������������桡����䡡������������������ۣʣݣ������������Σ�����������������������������������������������
�����ۣ��ݣӣ�����������ţ��ӣ�������㡡��������������������䡡����䡡����������ۣʣݣ��Σ������壬����������������������������������������������
�����ۣ��ݣɣ���������ţ��ţ��������졡���������������䡡��������������桡����������������������ۣʣݣ��У��������������У�������ţ���������ţ�������������䡡�ӣ��������������������������������������������
�����ۣ��ݣӣ���衡�����ǣ����������ȣ��ǣ�����顡�ͣ��������죮�У������㡡�����������������������������㡡�������죺��������������䡡������������������������ᣭ���塡������������ۣʣݣ��ţ�����������������ţ����������������������������������������
�����ۣ������ף�����������Ӣ�����������Լ�����̬������о���չ�ۣʣݣ�ֲ����̬ѧ������������������������������������������
�ۣ��ݣ£�����ʡ��ͣ��£�����á��ã��ǣ����������������������������ȣ����������ᡡ�������������������������������ۣʣݣ��£����������桡����塡�ԣ�������£��������졡�ã���⣬����������������������������������
�����ۣ��ݣ£��������ӣ��䡡�����������������䡡������������������������ӣ��������ᡡ���������ۣʣݣ��֣������������������������������������������
�����ۣ��ݣף�顡�٣��ģ��硡�ͣ��ȣ����硡�ڡ��٣��ӣ��䡡������������������������������䡡���������������桡�ӣ�����ᡡ�������ã������������壩���ᡡ������������������������������졡�����������硡�ʣ������£�����桡�أ������磬�ã�飭���ۣʣݣ���������������ʣ������졡��桡�£����������������������������������������
�����ۣ������������ͮ�����¹ڣ��ȣ��ֳ���ë�����������Լ����ȷ������о��ۣʣݣ�ʯ���Ӵ�ѧѧ������Ȼ��ѧ�棬��������������������������������������
�ۣ����ݣͣ�ţ�������С��£��ģ�������������䡡���������졡������������㡡�ᣭ�������桡���������������������ӣ�������������ᡡ�̣����ã���������壩�ۣʣݣ��ϣ�������ᣬ��������������������������������
�����ۣ�����Ҧ���죬�����ڣ��춰�����ȣ��½��ɺ���ֲ�������������Լ��� �ȷ����� �ۣʣݣ���̬ѧ����������������������������������������������
�����ۣ����ݣͣ����ᡡ���������ʣ����ʣ��ͣ���í��������á���������������죮�ͣ�������������������������䡡���������������������塡��桡���������������㡡��������桡����������������������ᡡ�ʣ��ǣ���������������壩���ᡡ������������������������䡡��������ۣʣݣ��ӣ��䡡�ӣ�飭����塡�ң������裬����������������������������������������
�����ۣ����ݣң�������ó��������ӣ��ͣ���Σ������ţ��ã��������é�ʣ������죮�ã�������������桡�������������졡�����������������塡��������������������������������㡡������桡�ӣ�����ᡡ����������ۣʣݣ���������桡�£������������������������������������������
�����ۣ����ݣԣ�������ᡡ�ͣ���������ᡡ�ͣ��ϣ������С��̣��ţ������������桡�������������������ᡡ���������������졡�������������ᡡ����������������硡�����������������㡡�ң��������������������������ᡡ�������ۣʣݣ��ʣ������졡��桡�ţ����������������������������������������������������
�����ۣ����ݣ̣�顡�ף��磬�ʣ�������͡��£�����ã���졡�á��£�����������죮�ӣ������������������������������䡡�����������������������������������������䡡�������������塡��������������������塡�ӣ�����ᡡ����������飭������ᡡ�����������塡����������졡���������ۣʣݣ��£ͣá��У�������£��������������������������������
�����ۣ�����ë�������ŵ����½������紺����ֲ����ϵ��Ҫ�ۣʣݣ��ɺ����о���������������������������������
�����ۣ�������ѧ�������������ȣ��½�ֲ�����ֶ����Ե��ص�����ۣʣݣ�ʯ���Ӵ�ѧѧ������Ȼ��ѧ�棬��������������������������������������
�����ۣ���������ɣ�κ�ң�ɢ֦��ë�˵����Ӷ����Լ����ȷ���Ϊ�ۣʣݣ�ֲ����̬ѧ�����������������������������������������������ۣ����������ᣬ�ź��������ֲ���ص���ֲ����ϵ�γɵ�̽�֣ۣʣݣ���ɽ��ѧѧ���۴ԣ�������������������������������
�ۣ����������ˣ��²��ƣ��۹Ŷ���ͨ����ɳĮֲ������Ե�һ���ص�ۣʣݣ���̬ѧ������������������������������������������������
�����ۣ����ݳ²��ƣ������ˣ����Ŀ����Ŷ���ͨ����ɳĮ��ɳ��ֲ��Ⱥ�䡢��ϵ����ֲ��Ļ��������ۣʣݣ�ֲ����̬ѧ���ֲ��ѧ�Կ�����������������������������������
�����ۣ����ݿ����أ����£���������½���������Į����ֲ��Ļ�ѧ�ɷֺ����ص�ۣʣݣ���̬ѧ����������������������������������������������
�����ۣ����������ƺ�Ƽ��Χ�������ɹŵ��Ͳ�ԭ���˻���ԭȺ��Ļָ������̼�ش�Ĺ��� �ۣʣݣ���Ȼ��ѧ��չ����������������������������������������
�����ۣ����ݼ���������ͮ���泻���ȣ��Ŷ���ͨ����ɳĮ��Ե��Ҫ��ľ�Ŀռ�ֲ���֣ۣʣݣ�ʯ���Ӵ�ѧѧ������Ȼ��ѧ�棬��������������������������������������
�����ۣ��������k�ģ��ҹ���Įֲ����ϵ�γɵ�̽�֣ۣʣݣ�ֲ�����ѧ������������������������������������������
�����ۣ����ݣȣ����ʡ��̣��̣�����졡�С��ȣ��ͣ���塡�ˡ��ǣ��ԣ�塡��������䡡��������桡������ۣʣݣ���������졡�ң����������桡�ţ���������ţ��������������ӣ������������������������������������������
�����ۣ����ݣգ���ɡ������У����������������������桡�����������塡������ۣʣݣ��£�������졡�ң����������������������������������������
�����ۣ����ݣԣ�������С������֣��������������䡡�����塡���������������ᣭ�������桡�ӣ����塡������ᣨ�̣����ã�������������������������������ۣʣݣ���������桡�£����������������������������������������