����ժ Ҫ������ԭø���к�ǿ�������ԣ��併�⽺ԭ�������ڶ���ϸ������ʵ����彵�������Źؼ����á����Ƚ��������コԭø�ķ���ͽṹ��������Ŀǰ���コԭø���Բⶨ����ԭø�����¡������ʹ��������ļ�����������Ȼ�������˽�ԭø��Ƥ�﹤ҵ��ҽҩ��ҵ��ϸ������������Ӧ�ü���ǰ�������Խ�ԭø�ִ��ڵ�����Ҳ���˼�Ҫ�ĸ���������δ���ķ�չ���������չ����
�����ؼ��ʣ�������; ��ԭø; ����; Ӧ��; �о���չ;
����Abstract����Collagenase is highly specific and plays a key role in the overall degradation of animal extracellular matrix due to its ability to degrade collagen. This review introduced the classification and structure of collagenase, and expounded the techniques and methods used in the activity determination, gene cloning, expression, purification and characterization of microbial collagenase. The application and prospect of collagenase was then summarized in leather industry, medical industry, cell separation and other fields. Finally, the existing problems for collagenase were briefly summarized, and the future development direction was prospected.
����Keyword����microorganism; collagenase; classification; application; research progress;
����ǰ��
������ԭ�����Ƕ�������֯����Ƥ�������������ǡ��졢Ѫ�ܣ�����Ҫ�ṹ���ס�������������Ҫ�ظ�Gly-X-Y�������������е�α����ɣ�X��Yͨ��������������Ǹ����ᣩ������ʹÿ�����������������������Եĵ����ļ��ṹ[1]����ԭ����ˮ��Զ�����������������DZز����ٵģ�������֯�ؽ�����̬�γɺ��˿����ϵ�[2]�����磺�������������������ϳɽ�ԭ��������½����ơ�����һЩ���岿λ��������ȴ�����ӣ���ԭ�����ľͿ���Ӧ����һ״������ԭ�������ǽ�ԭ����ˮ�������ջ���ҵ������ʳƷ����������кܸߵ����ü�ֵ�����⣬��ԭ��������ҽѧ��ҩѧ����Ҳ���ж���������ԣ��������ƹ�������֢��θ����Ѫѹ�ȶ��ּ�����Ҳ���������Ʊ�Ƥ���ı�ʪ��[3]����˽�ԭ����������ҵ�ϵ�Ӧ�ü�ֵ����Ѹ��������
����Ŀǰ����ԭ����ˮ�ⷽ�����ᷨ�����ø���������ᷨˮ��ͼˮ����ƻ������ᣬ����Щ������ȣ�ø��ˮ�ⷴӦ�ºͣ��������ƻ���С����ȫ�ɿ�������Ⱦ���������Ǿ��и߶ȵ������ԣ����н�ԭø���ǿ���������p H�����ͽӽ�������¶ȵ�����·������ã��ڼ��͵�Ũ�����Ծ��кܸߵĻ��ԡ����ſ�ѧ�����ķ�չ�����Ƿ��ֺܶ������ж����н�ԭø���ر�������ڶ�����Դ�Ľ�ԭø��˵�������ڱ���ʹ����������コԭø���ص�ˮ����ԣ�ʹ���������\����ҽҩ��ʳƷ�����졢����ӹ�����ױƷ���������ϵ����������Ź㷺��Ӧ��[4,5,6,7]��
����1 ����ԭ����
������ԭ����������߷��ӣ�Ҳ�Dz��鶯�����ں�����ࡢ�ֲ����Ĺ����Ե��ף�ռ���ڵ�����������25%��30%����ijЩ��������ռ�ı��������ߴ�80%[8]����ԭ�����ӳʰ�״����Լ300 nm��ֱ����1.5 nm���ң���Է�������Լ200��300 k Da����ԭ������3��α����ɣ�ÿ������Ҫ���ظ�������������Gly-X-Y������ɣ��Ǹ�����л����γ�����ȶ���ԭ�����ӣ���������ֱ��Ӱ���˽�ԭ������ά�����ȶ��Ժͻ�еǿ�ȡ�������α��ͨ����������۽��������»���������������ʽ���Ƴ����������ṹ��ʹ��ԭ�����кܸߵ�����ǿ��[9,10]��
������ԭ������Ϊ������Ͼ��кܵ͵�����ԭ�ԣ��ڲ������ĵ����������ԭ�Ը���[11]����ԭ���ױ����ǹ���ϸ������ʵĹǼܵ���Ҫ���ӣ������������ṹ���������γɵ���ά�������ϸ�������ȶ���֧�����ã���Ϊϸ������ֵ�����ṩ�ʵ��Ļ�����������������ϸ������֮֯�������õ������[12]����ԭ�����ܱ�����øˮ�⣬����������õ����オ����[13]����ԭ��������ֹѪ����[14,15]��ͨ���ٽ�ѪС�����ۺ�Ѫ����������Чʵ��ֹѪ����ԭ���������е���������ѧ���ܣ�ʹ����ҽѧ�����ݣ���ʳƷ��Ӫ��ѧ����Ƥ�ͨ�����Դٽ��Ƹ�����л����������գ�����������ʮ�ֹ㷺��Ӧ�á�
����2�� ��ԭø
������ԭøͨ������Ϊ��������ˮ����Ȼ��ԭ����ˮ���Ա��Խ�ԭ����ø���ӽ�ԭø��Դ�ĽǶȳ�������ԭøһ��ɷ�Ϊֲ�コԭø�����コԭø�����コԭø����ͼ1��ʾ�����ݱ������ӽ�������������ά��[16]���������齺[17]���Ѿ�����������˽�ԭø������ֲ�コԭø�������ض�λ���и���Ȼ��ԭ���ס����コԭøͨ���ֲ��ڼ��������ڣ����������Ȼ��ԭ��������α���ϵĵ����ļ�ˮ�⽺ԭ����X-Gly���Ϸ��ӵ��ķ�֮�������иԭ�����������ṹ��
����ͼ1 ��ԭø�ķ���
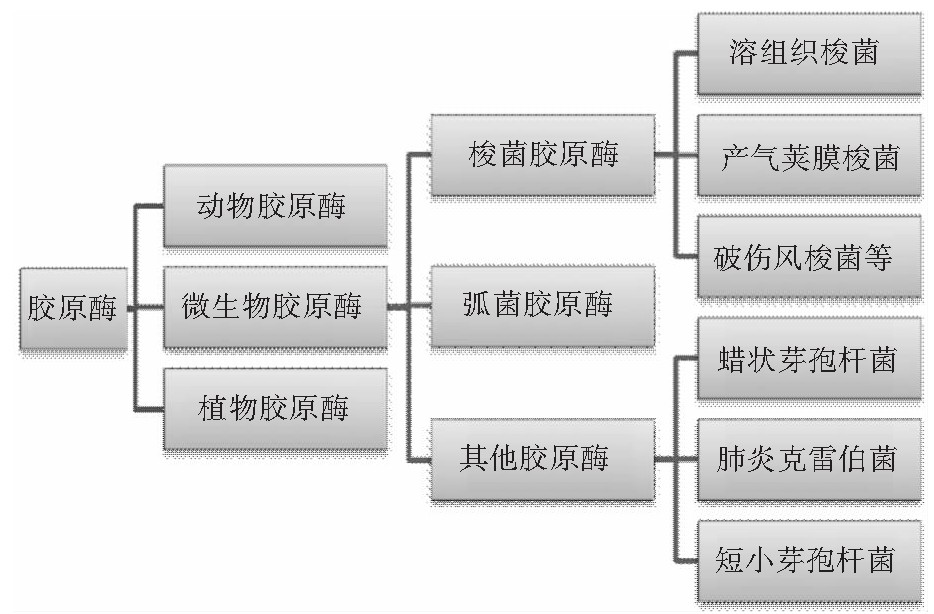
����Fig.1 Classification of microbial collagenase
�������コԭøʵ������һ�ֽ�������ø��������Zn2+��Ca2+�����ӷ��ӻ��ԡ�����������ˮ����ԵĿ����Խ�ԭ���ף����ҿ���ˮ����Ȼ�����Խ�ԭ����[18]���붯�コԭø��ȣ����コԭø����������㷺��ˮ���������ס��е����コԭø�����뽺ԭ���Ķ��λ�㷢����Ӧ��������С���ȵ�5���л��Ķ��ġ����Ƕ��コԭøֻ���뽺ԭ����N��3/4��Gly-Leu��Gly-lle�ļ�������Ӧ������һ��3/4�Ķκ�1/4�Ķ�[19]�����コԭø������ã�����Ϊ������Խ���ԭø���ڵ����⣬ͨ�����Ϳ��Դ��������������コԭø����Ҫ������֯��������ȡ���������ҳɱ��ܸߡ�
����3 �����コԭø�ķ���
�������コԭø����ø���ݿ�������M9���壬����ݰ��������кʹ����ܵIJ�ͬ����ΪM9A��M9B�����Ǽ��壬������Դ������ͻ����Ľ�ԭø���㷺�о�[20]��
����3.1 �������ԭø
���������ԭø����M9B�Ǽ��壬�����о����ֵ�����Դ������֯����Ľ�ԭø��1937�꣬��˹����Maschmann���״η��ֲ������˵�һ�������ԭø�����ѧ���ǽ����˸���������о���������Բ�ͬ�����ԭ���ף��������ϳɵ���ȣ�����Ի��ԡ��ȶ����Լ���ԭø��һ���������������ṹ����������֯�����ԭø����Ϊ���࣬��I���II�ེԭø[21]������I�ེԭø����Ȼ��ԭ���и�ˮ����ԣ���FALGPA�ϳɽ�ԭ�ľ����е�ˮ����ԣ���II�ེԭø����Ȼ��ԭ�����е�ˮ����ԣ���FALGPA�ϳ��ľ��и�ˮ����ԡ�Matsushita�о�С��������֯���Ⱦɫ���з�����������ԭø����col G��col H������I�ེԭø��col G�����룬��II�ེԭø��col H������[22,23]������֮�⣬��ѧ�һ�����������������ֽ�ԭø�������������磺������Ĥ�����col A����[24]�����˷������col T����[25]�ȡ�
��������֯�����ԭø������һ�ֶ�ṹ��ĵ����ʣ����ེԭøcol G��col H�ڽṹ��Ҳ�����Ų��졣col G��ԭø��S1��S2��S3a��S3b��4���ṹ����ɣ�col H��ԭø��S1��S2a��S2b��S3��4���ṹ�����[26]������S1�ǽ�ԭø�ṹ���еĴ��ṹ��(AD),S2��S3�ֱ����PKD��CBD�ṹ��PD����ø�ṹ����ͼ2��ʾ�����о�������col G��ԭø��S3b���н�Ͻ�ԭ�����������ṹ�����Dz�����ȱ���������������ĺ����������Խ�ԭ����ϡ�����col H��ԭø��S3�ṹ������col G��ԭø��S3b�ṹ���ƣ����ǹ���30%�İ��������У�S3b�в�������ϵ�9���л��У�ֻ��3������л���������S3�У��������жԱȣ�S3b�е�һ��Ca2+���춬���ᣨAsp927����S3�е�˿���ᣨSer896����ȡ��[27]��PKD�ṹ���CBD�ṹ�������������ԭ����ˮ�ⲻ�DZ���ģ����Ƕ��ڽϴ�Ľ�ԭʵ�壨���罺ԭ��ά��ȴ�DZ����[28]��I���II�ེԭø������λ���ǽ�ԭ����α�����ظ�Gly-X-Y�����е�Y-Gly���������о�֤�������ེԭø�ڽ�ԭ����ˮ���о���Эͬ����[29]��
����3.2�� ������ԭø
����������ԭø����M9A�Ǽ��塣�����������������ø����ΪI���II���������ø����I���������ø�У�п��ϻ������HEXXH-E���У���II���������ø����һ�µ�HEXXH���к�һ���Ȱ�����Ϊλ��HEXXH�������εĵ�����п�������[30]���������ݰ��������С�ø�������������������Ժ��ܣ������������������ø�����·�Ϊ����[31]����ͼ3��ʾ���������൰��ø���Ȼ�ĩ�˰���һ��PKD�ṹ���һ��PPC�ṹ��ǰ��ø�ṹ�����ڻ�������ø�е�PPC�ṹ��Ľṹ���ܻ���δ֪�ģ���Ҳ������ָ�������Դ��˻�����PPC�ṹ����IV�ͽ�ԭ���������[32]���������з����ͽṹ������֤�ݱ����������Եڶ��ࣨMEROPS M09.004���͵����ࣨMEROPS M09.001���Ļ�����������ø����Ϊ��ԭø[33,34]��������ԭøˮ�⽺ԭ�ĵ�һ���������и���������ԭ���ľ�Nĩ���ķ�֮��λ�õ�X-Gly�ļ��������Ƕ��コԭø��Gly-Leu��Gly-Ile�ļ�����ʵ�ϣ��������ԭøһ����������ԭø��Leu-Gly����ˮ��ϳ���Pz-Pro-Leu-Gly-Alu-D-Arg��
����ͼ2 colG��colH��ԭø�ṹͼ
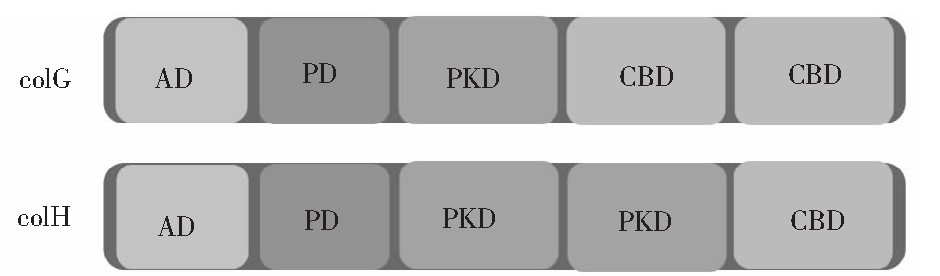
����Fig.2 The domain structures of col G and col H Collagenase
����ͼ3 �������������ø�ṹͼ
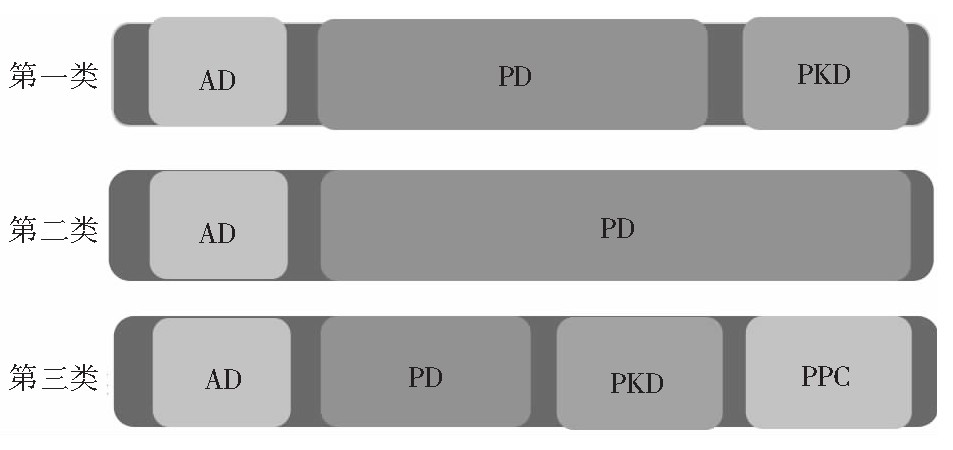
����Fig.3 The domain structures of three types of metallopro-teinases from vibrio
����3.3 ��������Դ
�����������ԭø�ͻ�����ԭø����Сѿ�߸˾�[36]������ѿ�߸˾�[37]���ݲ�ѿ�߸˾�[38]����ɫ�����[39]�ȶ��ɲ�����ԭø��������Щ���������ԭø�ľ���ṹ��δ��ȫ������������λ����δ��ȷ��Punta[40]���˷��������ڶ����コԭø�У������������Ľṹ������������������ͬ�������ֽṹ��Ľṹ���������ԡ�Abfalter����[41]������ѿ�߸˾�ATCC 14579 col A��ȫ��������������ΪĿ�����У�����BLAST�������бȶԵõ�����ѿ�߸˾���Y93-K850��col G����49.8%��ͬԴ��,�ʲ���SWISS-MODEL��col GΪָ��ģ���Y93-K850������ͬԴ��ģ�����ִ�ģ�Ͱ�������ṹ���ǽṹ��������ǽṹ����Щ���ֿ���ʹ���Ǹ�������˽����コԭø�Ĺ��ܺͻ��ƣ�����ȷ���ǵ�Ӧ�÷�����ˣ������コԭø������������и�����о��Ǻ��б�Ҫ�ġ�
����4 �����コԭø�Ļ��Բⶨ
�������コԭø���ڶҵ������кܸߵ�Ӧ�ü�ֵ����˲ⶨ��ԭø�Ļ��Ա��ʮ����Ҫ���������������ͪ��ɫ���ⶨ��ԭø�Ļ��ԣ���ˮ���Ի�ˮ�����Խ�ԭ����������ԭøˮ��ĵ���÷ֹ��ȼƲ������コԭ���ɵİ�����������ȷ����ԭø���ԡ��˷���һ�㽫1min�����ض���������ˮ���������൱��1μg�������ø������Ϊ1��ø������λ������֮�⣬�ϳ���N-(3-[2-fury]acryloyo)-Leu-Gly-Pro-Ala(FALGPA��Ҳ������Ϊ��Ч�ⶨ��ԭø���Եĵ����������������ﷴӦ��ΪѸ��[42]��Zhang���˻�����14C,2H��3H�����Ա�ǻ�ӫ���ǵĽ�ԭ��Ϊ���������������ԭø����[43]������ͪ��ɫ�����ܻ�������Ӱ�죬���ϳ��ĺ�ӫ���ǽ�ԭ������Ϊ������Ҫ�߳ɱ������·����Է���IJ������������ڶ�ⶨ���У���Ȼû��һ�ּȼɿ��־��õķ����ⶨ��ԭø���ԡ�
����5 �����コԭø�Ļ����¡�������ͱ���
��������Ŀ�¡�ͺ��������м��Ϊ����ø�Ľṹ�����ṩ�˰����ͱ��������ڣ���ѧ���Ѿ���һЩ�����д����������コԭø�������Ѿ���¡�����ǵ���Ӧ����������˲�������֯�����col G[44]��col H[45]�����˷�˾���col T����[45]���ѱ��ɹ���¡�����ڴ˾������������ϻ����Ľ�ԭø����Ҳ�ѱ���¡����ȷ��������������������[46]�����コԭø�����ķ���������麟��������ˡ������������������˲��������ӽ���������[47]�������ڴ����ɱ��ܸߣ�Ҳ�����������コԭø��һЩ��ҵ�����Ӧ�á�
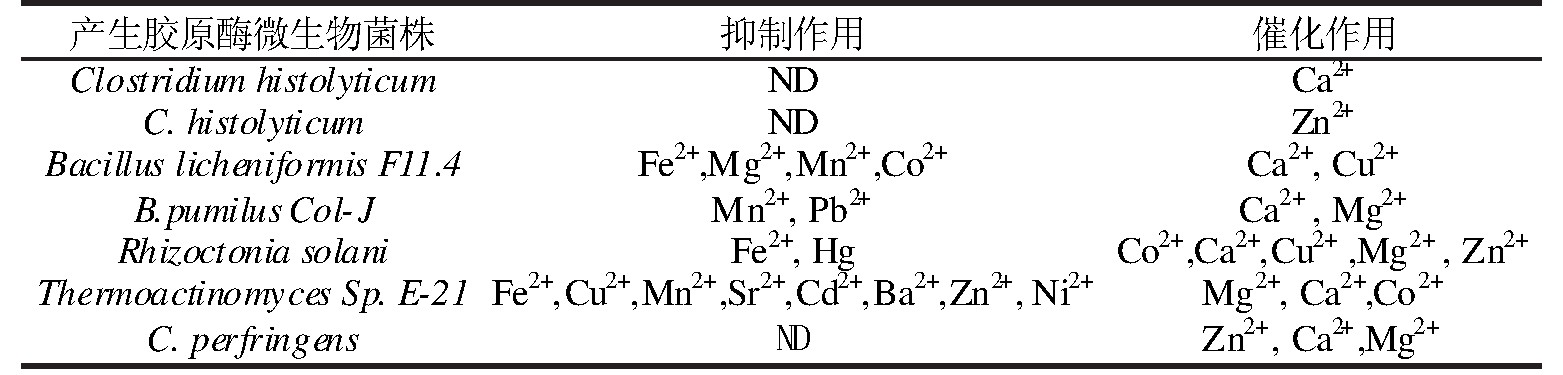
�������コԭø�ı������Դ����ǵķ�������������ѧ�����Լ�p H�����ȶ��ԡ��������Ӷ�ø������Ӱ��ȷ������֡��ⶨø���������ķ�����ʮ����������ƾ۱�ϩ����������Ӿ����������ɫ������ЧҺ��ɫ����������������ü����ȣ��ı����Ũ��ʱ����Ӧ���ʻ���֮�仯����ʱ������˫������ͼ������ó�����ѧ���������コԭø�ڷ�Ӧ�е�����p H�������¶Ⱥ�����Ӱ�����ؿ���ͨ���ⶨø������ȷ�������н��������ڸ������コԭø�����о�����Ҫ���ã��е�������ٽ����ã���Ca2+��Zn2+�����е����ӻ����������ã���Fe2+��Mn2+[31]�����1��ʾ��
������1 ��ͬ���ӶԲ�ͬ��Դ��ԭø���Ե�Ӱ��
����ND=not determined
����6�� ���コԭø��Ӧ��
����6.1�� Ƥ�﹤�յ�Ӧ��ǰ��
������ԭø��Ƥ�﹤ҵ���кܶ�DZ�ڵ�Ӧ��ǰ����Ƥ�﹤ҵ�н�ˮ����֬����ë���ҡ��ѻҡ����������ᡢ���ơ��к͡�������Ⱦɫ��֬������������ʹ�û������Ϸ��ӣ��Ի����кฺܶ��Ӱ�졣Ƥ���ҵ�����ſ���������Ч�Ƹ﹤�յľ�ѹ�����Դ������ִ���Ⱦ���ŷŷ����Ҫ�����Ѱ��Ƥ���Ƥ�ﻯѧ�ӹ��еĻ������Ʒ��Ƥ�﹤ҵ���ٵľ���ս�����ⷽ�棬��Ϊ��ѧ�ӹ��Լ����Ʒ��ø�����ջ����һ�µ��Ƽ�����Ϊ���Dz������������������õIJ�Ʒ�����Ҽ�����Σ�պ���Ⱦ��ѧƷ��ʹ��[48]��Ⱦɫ��Ƥ�﹤ҵ��һ����Ҫ���գ���Ƥ�﹤ҵ��ˮ�д��ڵ�δ�����Ⱦ���������������п��ԣ�����һ����Ҫ���⡣Ƥ�����ƺ����Ƿ���ʹ�����コԭø�ᵼ����ά״Ƥ������Ŀ��ţ��ٽ�Ⱦ���ڻ����е���ɢ��ʹ������ø�����ڱ�֤Ƥ��ǿ�Ȳ���������ʹƤ�������������Ⱦ�ϵ������ʿɴ�99%��ͬʱҲ������Ƥ��ķ����ȡ�����ƽ���ȡ��ָк���ۣ��������DZ������������������Ⱦ�ϵ�Ⱦɫ����[49]��������Ƥ���ˮ��Ⱦ�ϵ��ŷš������Ƹ�����е���ë����Ҳ�Ի�����������ص���Ⱦ�����Ѱ����̬�Ѻõ����Ʒ�Ǻ��б�Ҫ�ġ�Sivasubramanian����[50]����˽����コԭø������ë���գ������ڼ�����Ⱦ���ɺͽ�ԭ�������ˡ�Ȼ�������コԭø�������ɱ���������������������ë�����е���ҵӦ�á�
����6.2�� ҽҩ��ҵ��Ӧ��
�������コԭø��ҽҩ��ҵ�о�����Ҫ��λ����ԭ����ռ���嵰���ʵ�����֮һ����ӳ�����ڽ����ͼ����е��������á����コԭø��ֱ��Ӧ�����ٴ����ƣ��������崴[51]�������[52]��������ͻ��[53]�ȡ�
�����ƽ�Ĥ����֢��DD������һ�ֽ����Է�չ����ά����ֳ�Լ�������Ҫ���ƽ�Ĥ�������ָ��Ĥ�����������ָ�ؽں�ָ��ؽڵĹ������ϰ�������������ø��DD�����е��״�Ӧ������1965��[54]����ʱʹ�õ����ȵ���ø��������ø�Ļ���������ȷ�˳����ĵ�����δ����Ģ��ͽ�ԭ����ʹ�ý�ԭø�Ƚ�Ĥ�г����������㣬��Ϊ���Ǿ��н�ԭ�����Եġ�2010������ʳƷҩƷ�ල�����֣�FDA��������֯�����ԭø��������DD�������ơ��ھ���Ӧ��ʱ��ϸ��ˮƽ�ϣ����ƽ�ԭø����������ʱ��ȥ���ƻ����Ʋ�����άϸ������ɢ�����ź���ֳ��ת¼ˮƽ�ϣ����ƽ�ԭø�����ɶԶ���ECM�ɷ֡�ϸ�����Ӻ��������ӽ�������[55]��Badalamente����[56]�ϸ���ѭҪ��ѡ����35�����껼�ߣ��еĻ���MP����ָ�ؽڣ������̶���20°��100°֮�䣬�еĻ���PIP������ָ��ؽڣ������̶���20°��80°֮�䣬�ų�ʹ���Ļ��أ���ԭø���������أ�����ѡ��Ļ���ע�佺ԭø�����ע��3�Σ��������ټ�¼������21�����ߵĻָ����̣��������̶Ȼָ���5°��Ϊ��㣬ע��1��24���º�86%MP���ߺ�100%PIP���߶�����Ч���Ƚ����⣬�Ҹ���CMS���ִﵽ79�ֿ���˵����ԭø��DD��������Ч�����á�
������һЩ�о��У�������Ϊ���ܽ�ԭø���ƺͽ�Ĥ�п���û�в��죬������Ƚ�Ĥ�п�������ԭø�����Ƴɱ��ϵ�[57]��Naam[58]���о����������ʹ�ý�ԭø���Ƶ�DD���ָ��������ճ��ʱ�����ʹ���������Ƶ�ʱ��Ҫ��һЩ����ԭø��Ա������飺1.9 d�Ա�37.4 d������������౨���о�֤�������ý�ԭø����DD�������ʽϵͣ����ز����¼������������ˣ�����ע�佺ԭø�ķ�����DD���߽��������ǿ��еģ�Ӧ���Թ�����
����6.3�� ϸ�����뷽���Ӧ��
����ϸ������һ����ԭ��������ϸ�������ϸ��ϵ�ȼ����������ͣ�����ø���������ڴ��������б�������ϸ����������֯�ֽ��ϸ������[59]���ȵ���øͨ�����ڲ��鶯��ϸ������֯����ͷ��룬��ʵ���б��㷺��������������������������ϵ�ϸ���ķ��롣����ϸ��������������Ҫ�����ǵ�ϸ���ﵽ�߶��ں�ʱ��ϸ���ӻ����з������[60]��Ȼ�����ȵ���ø��������;�����л����Ȼ������������ѽ�[61]���䵰��ˮ����Կ���ͨ���ѽ�ϸ�������������������Ĥ��������ϸ������ˣ�Ϊ�˻���������Ļϸ����ѡ���ʵ���ø��������Ҫ�ġ���һ���棬��ԭ�����ߵ���ά��֯���ȵ���øˮ����еֿ�������������Щ��֯��ʹ���ȵ���ø����Ч���к���[62]��
�������コԭø��ϸ����ά��֯��������֯��������к��м�ֵ[63]��Sun����[64]ͨ��0.01%���コԭø��33%Percoll�ܶ��ݶ����Ĵ�С������������Ʒ���������ʹ�������ϸ����ʹ����и��ߵ�ϸ�������ͻ�����Tanaka����[65]����˽��ɴ��ṹ����ɵ�����62 k Da��ԭø����ϸ�����룬���кܸߵ�����Ч�ʣ���֤�����������齺ԭø���������������ȶ��ͻ�Ծ�ģ���������������֯����ı���ԭø��Ʒ��ȣ������и��ߵ�ˮ�⽺ԭ�����ԣ������и��������Ľ�ԭ���ס�
����7 ��������
����Ŀǰ����ѧ����ڽ�ԭø�ĸ������ʶ��塢����ͼ����������ž�����顣����ø�ı����ǵ����ʣ����Խ�ԭø�ڱ���ʵ��Ͳ��뷴Ӧ�Ĺ����������¶ȡ�p H��������Ӱ�죬����ȡ���コԭø�Ĺ����У�Ҳ���кܶั���ͬʱ��Ϊ�ɱ���Ҳ����ʵ�ֹ�ҵ�����������コԭø��Ӧ���ϣ�ҽѧ����Ӧ�������ٴ���⣬���ٵ��в��˵Ļָ���������ʣ���ʳƷ������ҪѰ�ҷ��²������в������齺ԭø���䴿�������ⲡ�����ڷ�����Щø��ʳƷ��ҵ�еİ�ȫʹ�ã�����δ�����о�Ӧ�ü�����������Խ�ԭ���ġ���ԭˮ�������������������ʳƷ�Ŀ����ϣ�Ƥ��������Ҫ����߲�Ʒ������ͬʱ��Ҫ���ӻ�������̬���⣬���÷ǻ�ѧ��ʽ���������������ʽ����Ƥ���ҵ����̬������ˣ�ɸѡ��Ч�IJ���ԭø���������Ϳ����ͳɱ��������Լ�ͨ����ͬ�������������õ��������������ԵĽ�ԭø������Ƥ���ҵ����̬���ͳ�����չҲ������ʮ����Ҫ�����塣
���������
����[1] Adhikari A S,Glassey E,Dunn A R.Conformational dynamics accompanying the proteolytic degradation of trimeric collagen I by Collagenases[J].Journal of the American Chemical Society,2012,134(32):13259-13265.
����[2] Bilek S E,Bayram S K.Fruit juice drink production containing hydrolyzed collagen[J].Journal of Functional Foods,2015,14:562-569.
����[3] Lima C A,Freitas A C V,Lima J L,et al.Two-phase partitioning and partial characterization of a Collagenase from Penicillium aurantiogriseum URM4622:Application to collagen hydrolysis[J].Biochemical Engineering Journal,2013,75:64-71.
����[4] Daboor S M,Budge S M,Ghaly A E,et al.Isolation and activation of Collagenase from fish processing waste[J].Advances in Bioscience and Biotechnology,2012,3 (3):191-203.
����[5] Daboor S M,Budge S M,Ghaly A E,et al.Extraction and purification of Collagenase enzymes:A critical review[J].American Journal of Biochemistry and Biotechnology,2010,6(4):239-263.
����[6] Jain R,Jain P C.Production and partial characterization of Collagenase of Streptomyces exfoliatus CFS 1068 using poultry feather[J].Indian Journal of Experimental Biology,2010,48(2):174-178.
����[7] Kaur S P,Azmi W.Cost effective production of a novel Collagenase from a non-pathogenic isolate Bacillus tequilensis[J].Current Biotechnology,2013,2(1):17-22.
����[8] Zuber M,Zia F,Zia K M,et al.Collagen based polyurethanes-A review of recent advances and perspective[J].International Journal of Biological Macromolecules,2015,80:366-374.
����[9] ����.�ǻ������ζԽ�ԭ���ȶ���Ӱ���о�[D].ɽ��ũҵ��ѧ��2015.
����[10] Gonzalez E A,Bielajew B J,Hu J C,et al.Engineering self-assembled neomenisci through combination of matrix augmentation and directional remodelin[J].Acta Biomaterialia,2020,109:73-81.
����[11] Gharenaz N M,Movahedin M,Mazaheri Z.Three-dimensional culture of mouse spermatogonial stem cells using a decellularised testicular scaffold[J].Cell Journal,2020,21(4):410-418.
����[12] Yuan J J,Zhou Q,Wang P,et al.Phosphorylation of silk fibroin via maillard reaction and its behavior of biomimetic mineralization[J].Fibers and Polymers,2019,20 (8):1616-1623.
����[13] Jiang X J,Wang Y,Fan D C,et al.A novel human-like collagen hemostatic sponge with uniform morphology,good biodegradability and biocompatibility[J].Journal of Biomaterials Applications,2017,31(8):1099-1107.
����[14] Slezak P,Monforte X,Ferguson J,et al.Properties of collagen-based hemostatic patch compared to oxidized cellulose-based patch[J].Journal of Materials Science-Materials in Medicine,2018,29(6):71.
����[15] Yuan H B,Chen L,Hong F F.A biodegradable antibacterial nanocomposite based on oxidized bacterial nanocellulose for rapid hemostasis and wound healing[J].ACS Applied Materials&Interfaces,2020,12(3):3382-3392.
����[16] Kim M,Hamilton S E,Guddat L W,et al.Plant Collagenase:unique colla enolytic activity of cysteine proteases from ginger[J].Biochimica Et Biophysica Acta-General Subjects,2007,1770(12):1627-1635.
����[17] Raskovic B,Bozovic O,Prodanovic R,et al.Identification,purification and characterization of a novel collagenolytic serine protease from fig (Ficus carica var.Brown Turkey) latex[J].Journal of Bioscience&Bioengineering,2014,118(6):622-627.
����[18] Santra M,Luthra-Guptasarma M.Assaying Collagenase activity by specific labeling of freshly generated N-termini with fluorescamine at mildly acidic p H[J].International Journal of Peptide Research and Therapeutics,2020,26(2):775-781.
����[19] Gomez D E,Alonso D F,Yoshiji H,et al.Tissue inhibitors of Metalloproteinases:structure,regulation and biological functions[J].European Journal of Cell Biology,1997,74 (2):111-122.
����[20] Rawlings N D,Barrett A J,Thomas P D,et al.The MEROPSdatabase of proteolytic enzymes,their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database[J].Nucleic Acids Research,2018,46(D1):D624-D632.
����[21] Duarte A S,Correia A,Esteves A C.Bacterial Collagenases-A review[J].Critical Reviews in Microbiology,2016,42(1):106-126.
����[22] Matsushita O,Jung C M,Katayama S,et al.Gene duplication and multiplicity of Collagenases in Clostridium histolyticum[J].Journal of Bacteriology,1999,181(3):923-933.
����[23] Zhang Y Z,Ran L Y,Li C Y,et al.Diversity,structures and collagen-degrading mechanisms of bacterial collagenolytic proteases:An overview[J].Applied&Environmental Microbiology,2015,81(18):6098-6107.
����[24] Obana N,Nomura N,Nakamura K.Structural requirement in Clostridium perfringens Collagenase m RNA 5'leader sequence for translational induction through small RNA-m R-NA base pairing[J].Journal of Bacteriology,2013,195(12):2937-2946.
����[25] Ducka P,Eckhard U,Sch?nauer E,et al.A universal strategy for high-yield production of soluble and functional clostridial Collagenases in E.coli[J].Applied Microbiology and Biotechnology,2009,83(6):1055-1065.
����[26] Sekiguchi H,Uchida K,Matsushita O,et al.Basic fibroblast growth factor fused with tandem collagen-binding domains from Clostridium histolyticum Collagenase col G increases bone formation[J].Biomed Research International,2018,2018:8393194.
����[27] Kumar P G,Nidheesh T,Suresh P V.Comparative study on characteristics and in vitro fibril formation ability of acid and pepsin soluble collagen from the skin of catla (Catla catla)and rohu (Labeo rohita)[J].Food Research International,2015,76(3):804-812.
����[28] Bauer R,Wilson J J,Philominathan S T L,et al.Structural comparison of col H and col G collagen-binding domains from Clostridium histolyticum[J].Journal of Bacteriology,2013,195(2):318-327.
����[29] Eckhard U,Brandstetter H.Polycystic kidney disease-like domains of clostridial Collagenases and their role in collagen recruitment[J].Biological Chemistry,2011,392 (11):1039-1045.
����[30] Rawlings N D,Waller M,Barrett A J,et al.Merops:the database of proteolytic enzymes their substrates and inhibitors[J].Nucleic Acids Reseaech 2014,42 (D1):D503-D509.
����[31] Pal G K,Pv S.Microbial Collagenases:challenges and prospects in production and potential applications in food and nutrition[J].RSC Advances,2016,6(40):33763-33780.
����[32] Miyoshi S.Vibrio vulnifificus infection and Metalloprotease[J].Indian Journal of Dermatology,2006,33:589-595.
����[33] Kim S K,Yang J Y,Cha J.Cloning and sequence analysis of a novel Metalloprotease gene from Vibrio parahaemolyticus04[J].Gene,2002,283(1-2):277-286.
����[34] Bauer R,Janowska K,Taylor K,et al.Supporting information for article:Structures of three polycystic kidney disease-like domains from Clostridium histolyticum Collagenase col G and col H[J].Acta Crystallographica.Section D:Biological Crystallography,2015,71:565-577.
����[35] Thring T S A,Hili P,Naughton D P.Anti-Collagenase,anti-elastase and anti-oxidant activities of extracts from 21plants[J].BMC Complementary and Alternative Medicine,2009,9:27.
����[36] Wu Q,Li C,Li C Y,et al.Purification and characterization of a novel Collagenase from Bacillus pumilus Col-J[J].Applied Biochemistry and Biotechnology,2010,160(1):129-139.
����[37] Chen S H,Ma M H,Fu X.Analyzing structural and functional characteristics of Collagenase from Bacillus cereus MH19 via in silico approaches[J].Current Proteomics,2020,17(3):200-212.
����[38] Tran L H,Nagano H.Isolation and characteristics of Bacillus subtilis CN2and its Collagenase production[J].Journal of Food Science,2002,67(3):1184-1187.
����[39] Lima C A,Rodrigues P M B,Porto T S,et al.Production of a Collagenase from Candida albicans URM3622[J].Biochemical Engineering Journal,2009,43(3):315-320.
����[40] Punta M,Coggill P C,Eberhardt R Y,et al.The Pfam protein families database[J].Nucleic Acids Reseacch,2012,40(D1):D290-D301.
����[41] Abfalter C M,Schoenauer E,Ponnuraj K,et al.Cloning,purification and characterization of the Collagenase col A expressed by Bacillus cereus ATCC 14579[J].Plos One,2016,11(9):e0162433.
����[42] Yoshida E,Noda H.Isolation and characterization of Collagenases I and II from Clostridium histolyticum[J].Biochimica Et Biophysica Acta,1965,105(3):562-574.
����[43] Zhang Y F,Fu Y,Zhou S F,et al.A straightforward ninhydrin-based method for Collagenase activity and inhibitor screening of Collagenase using spectrophotometry[J].Analytical Biochemistry,2013,437(1):46-48.
����[44] Eckhard U,Schoenauer E,Nüss D,et al.Structure of Collagenase G reveals a chew-and-digest mechanism of bacterial collagenolysis[J].Nature Structural&Molecular Biology,2011,18(10):1109-1114.
����[45] Ducka P,Eckhard U,Sch�inauer E,et al.A universal strategy for high-yield production of soluble and functional clostridial Collagenases in E.coli[J].Applied Microbiology and Biotechnology,2009,83(6):1055-1065.
����[46] Ohbayashi N,Matsumoto T,Shima H,et al.Solution structure of lostridial Collagenase H and its calcium-dependent global conformation change[J].Biophysical Journal,2013,104(7):1538-1545.
����[47] Eckhard U,Huesgen P F,Brandstetter H,et al.Proteomic protease specificity profiling of clostridial Collagenases reveals their intrinsic nature as dedicated degraders of collagen[J].Journal of Proteomics,2014,100:102-114.
����[48] Kaushik R,Saran S,Isar J,et al.Statistical optimization of medium components and growth conditions by response surface methodology to enhance lipase production by Aspergillus carneus[J].Journal of Molecular Catalysis B-Enzymatic,2006,40(3-4):121-126.
����[49] Kanth S V,Venba R,Madhan B,et al.Studies on the influence of bacterial Collagenase in leather dyeing[J].Dyes&Pigments,2008,76(2):338-347.
����[50] Sivasubramanian S,Manohar B M,Rajaram A,et al.E-cofriendly lime and sulfide free enzymatic dehairing of skins and hides using a bacterial alkaline protease[J].Chemosphere,2008,70(6):1015-1024.
����[51] Pham C H,Collier Z J,Fang M,et al.The role of Collagenase ointment in acute burns:a systematic review and meta-analysis[J].Journal of Wound Care,2019,28(2):S9-S15.
����[52] Song M,Lee S,Choe D,et al.Clinical and biological evaluations of biodegradable collagen matrices for glaucoma drainage device implantation[J].Investigative Opthalmology&Visual Science,2017,58(12):5329-5335.
����[53] Changhuai L U,Zhi J L,Hong B Z,et al.Mechanism of p38mitogen activated protein kinase signaling pathway on promoting the hypertrophy of human lumbar ligamentum flavum via transforming growth factorβ1/connective tissue growth factor[J].Chinese Journal of Reparative and Reconstructive surgery,2019,33(6):330-335.
����[54] Bassot J.Treatment of Dupuytren’s����ת��27ҳ��disease by isolated pharmacodynamic“exeresis”or“exseresis”completed by a solely cutaneous plastic step[J].Lille Chirurgical,1965,20:38-44.
����[55] Warwick D,Arandes-Renu J M,Pajardi G,et al.Collagenase Clostridium histolyticum:emerging practice patterns and treatment advances[J].Journal of Plastic Surgery and Hand Surgery,2016,50(5):251-261.
����[56] Badalamente M A,Hurst LC.Efficacy and safety of injectable mixed Collagenase subtypes in the treatment of Dupuytren's Contracture[J].Journal of Hand Surgery-American Volume,2007,32A(6):767-774.
����[57] Nydick J A,Olliff B W,Garcia M J,et al.A comparison of percutaneous needle fasciotomy and Collagenase injection for Dupuytren Disease[J].Journal of Hand Surgery-American Volume,2013,38A(12):2377-2380.
����[58] Naam N H.Functional outcome of Collagenase injections compared with fasciectomy in treatment of Dupuytren’s contracture[J].Hand (New York,N.Y.),2013,8 (4):410-416.
����[59] Duarte A S,Rosa N,Duarte E P,et al.Cardosins:a new and efficient plant enzymatic tool to dissociate neuronal cells for the establishment of cell cultures[J].Biotechnology and Bio-���Ͻӵ�22ҳ��engineering 2007,97(4):991-996.
����[60] Huang H L,Hsing H W,Lai T C,et al.Trypsin-induced proteome alteration during cell subculture in mammalian cells[J].Journal of Biomedical Science,2010,17:36.
����[61] Olsen J V,Ong S E,Mann M.Trypsin cleaves exclusively C-terminal to arginine and lysine residues[J].Molecular&Cellular Proteomics,2004,3(6):608-614.
����[62] Yu Z X,Visse R,Inouye M,et al.Defining requirements for Collagenase cleavage in collagen type III using a bacterial collagen system[J].Journal of Biological Chemistry,2012,287(27):22988-22997.
����[63] Santra M,Luthra G M.Assaying Collagenase activity by specific labeling of freshly generated N-Termini with fluorescamine at mildly acidi p H[J].Inrernational Journal of Peptide Research and Therapeutic,2020,26(2):775-781.
����[64] Sun C,Luo Q B,Lu X X,et al.Isolation and purification of primary Kupffer cells from mouse liver[J].Chinese Journal of Cellular&Molecular Immunology,2016,32(8):1021-1025
����[65] Tanaka K,Okitsu T,Teramura N,et al.Recombinant Collagenase from Grimontia hollisae as a tissue dissociation enzyme for isolating primary cells[J].Scientific Reports,2020,10(1):3927.





