
根据引起弱视的病因,常见的弱视可分为:斜视型弱视、屈光不正型弱视、屈光参差型弱视及形觉剥夺型弱视,其中以形觉剥夺型弱视治疗效果最差.
弱视的发病机制十分复杂,目前弱视发病机制的分子神经生物学研究方面取得了一些进展,尤其弱视发展中各级神经元间突触连接的可塑性变化的理论为治疗效果差的形觉剥夺性弱视提供了新的治疗契机.
基于此,本工作使用氟西汀粉剂,配置合适浓度,使用新生猫制作形觉剥夺型弱视模型,采用视觉诱发电位检测,观察视觉诱发电位(P- VEP)峰的潜伏期和波幅的变化,来评估氟西汀治疗形觉剥夺弱视模型猫的疗效.
材料和方法
实验动物及分组 健康清洁级新生猫(出生2 周,性别不限)18 只和母猫 3 只,新生猫出生时常规体检无异常眼疾;随机分为正常对照组 3 只,形觉剥夺组猫(5 组,每组 3 只,共15只).
建模:眼睑缝合操作[1]:自内眦到外眦剪除上下睑缘1.5mm,皮肤及皮下行分层水平褥式缝合,封闭睑裂,点左氧氟沙星眼液,每日3 次,共 7 天.正常对照组:不行眼睑缝合.
幼猫均在正常同等环境下散养,在进行视觉诱发电位前剪开缝合眼,并再次点左氧氟沙星眼液 3 次.所有幼猫用美多丽眼液扩大瞳孔,行视网膜检影,矫正屈光.12周后形觉剥夺弱视猫模型建成,开始使用氟西汀(Fluoxetine)药粉(浓度:20μmol/l,2mg/kg)给猫灌胃,每天 1次,连续 4 周.
视觉诱发电位检测采用德国 Roland 公司生产的 RETIScan3.20 视觉电生理仪,P- VEP波形图中的 P 波,及其前、后出现的 2 个的N1 和 N2 波,主要以 P 波的潜伏期及波幅为分析指标.分别在使用氟西汀 4 周后、8 周后、12 周后、16 周后、20 周后行猫的视觉诱发电位检测.P- VEP记录[1]:幼猫经氯氨酮(15mg/kg)腹腔麻醉,剃净幼猫额部、枕部及右耳背毛,在电屏蔽暗室内进行实验.采用自制钢针电极,将引导电极插入枕骨中线凹陷正上方 2cm 处颅骨下,参考电极置于鼻根部上方 lcm 处皮下,接地电极为右耳尖,单极引导.作用电极与参考电极间阻抗<5KΩ;采样时间 250ms,波形进行 128 次叠加平均.
数据分析:分析时用 RETIScan 3.20 自带程序分析 N1P1,N2P2 波的潜伏期及波幅.有关数据用 RETIScan 3.20 自带程序分析处理后,将初步分析结果输入 Excel,统计分析采用SPSS ll.0 软件.采用 t 检验,以 P < 0.05 为有统计学意义.
结 果
正常猫 P- VEP 分析(表 1)
正常猫单眼 P- VEP,由 1 个明显的正波 P波和其前较小的负波 N 波组成,P 波波幅均大于 N 波. P 波的波形及出现的时间较恒定,N波变异较 P 波大,通常记录不到 N2 波.本实验正常对照组PVEP 定量分析表明,P 波的平均 潜 伏 期 为 48.33 ±3.51ms, 平 均 波 幅 为55.69±3.52μV.

单眼剥夺猫 P- VEP 分析(表 1)
本实验组单眼形觉剥夺猫 P- VEP 定量分析 表 明 ,P波 的 潜 伏 期 延 长 , 平 均 值 为58.23±4.62ms,波幅增加,平均值为 38.24±2.96μV.组间比有统计学意义(P < 0.05).喂氟西汀治疗后单眼剥夺猫图形视觉诱发电位分析(表 2、表 3).
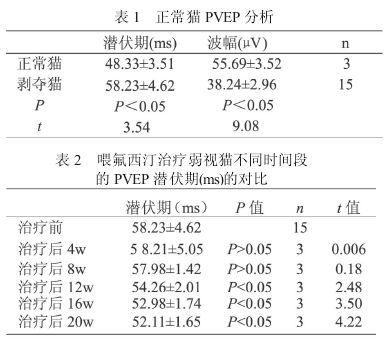
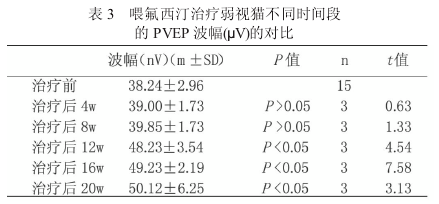
治疗 4 周、8 周后,P 波的潜伏期稍缩短,波幅稍增加,组间比较无统计学意义.治疗12周后,P 波潜伏期缩短,波幅增加,与治疗前、后组之间相比有统计学意义(P < 0.05),但仍低于正常对照组猫.治疗 16 周、20 周后,P波潜伏期进一步缩短及波幅进一步增加,与未经治疗组相比有统计学意义(P < 0.05,n=3).
讨 论
弱视是常见眼病,患病率约为 3%,我国目前人口 14 亿,儿童弱视患者估计达 3 千万,为数相当可观.若弱视治疗未愈,对这些儿童将来的学习和求职生涯产生巨大的影响,对这些家庭造成不必要的压力.在弱视的发病机制研究中,主要有"中枢病因学说"与"外周病因学说"."中枢病因学说"认为,弱视患者的主要受损部位在视皮层.
由于各种原因使视觉信息传入减少,致视皮层双眼性神经元发育障碍,视功能发育延迟而发生弱视."外周病因学说"认为,弱视是由于视网膜X型神经节细胞未受到足够刺激所致.两种学说相互联系,互为补充."中枢病因学说"的直接的电生理依据是弱视眼的P- VEP 以及多导 P- VEP 的 P 波振幅明显下降,伴或不伴随潜伏期的延长[2]."外周发生学说"的电生理依据是弱视眼仅损害了对高空间频率刺激的反应能力[3].
单眼睑缝合是研究发育期形觉剥夺效应的一种有价值的实验方法,能够藉此研究弱视动物模型的视觉电生理特征.弱视病理生理研究提示: 弱视眼 P- VEP 的异常源自视皮层对图形运动感觉及边界对比效应敏感的神经元的功能异常.但在单眼形觉剥夺猫的健眼,其P- VEP 的 P 波潜伏时与振幅随刺激空间频率的变化关系并不明显,Sclarll 等认为即使经长期的形觉剥夺后,皮层残留的抑制性突触连接仍可保持其形态和功能的完整性.本课题组采用德国Roland公司生产的 RETIS-can3.20 视觉电生理仪记录人类视觉诱发电位一般有的 N1、P1、N2 三个波,其中 P100潜伏期一般为 95~105ms,波幅 10~15μV.但本实验记录到的正常对照组猫的 P- VEP 与人类的不同,一般只能采集到 N1、P1 两个波,P 波的潜伏期平均为48.33 ±3.51ms,波幅平均55.69±3.52μV.
本课题组收集临床弱视患儿,发现其P- VEP出现潜伏期延长及波幅降低[4].本实验在单眼形觉剥夺模型猫记录到的剥夺眼P- VEP与人类弱视眼 P- VEP 的变化具有相同趋势.P 波潜伏期平均为 58.23±4.62ms,较正常对照组猫延长9.9ms,与正常对照组相比有统计学意义.
波幅平均 38.24±2.96μV,较正常对照组猫降低 17.45μV,也有统计学意义.这些实验结果表明,在视觉发育关键期行单眼睑缝合,剥夺形觉后,确实成功塑造出弱视动物模型.形态学与生理学的研究已经证实[5],单眼形觉剥夺的效应主要发生在视中枢.
视觉系统的正常发育明显受环境因素的影响.氟西汀作为选择性5- HT再摄取抑制剂(Selective sero-tonin reuptake inhibitor,SSRI)其作用机制是阻断 5- HT的再摄取,使神经突触间隙中可供生物利用的 5- HT 增多,从而增强 5- HT 能神经元传递作用.SSRI 通过使位于 5- HT神经末梢突触前膜上的 5- HT 自身受体敏感性降低,从而使每次神经冲动时释放的 5- HT 量增多.事实上,5- HT 是一部分兴奋性中间神经元的主要神经递质,5- HT 还可以调节或促进目的性运动反射.长期应用氟西汀不仅可上调 5- HT受体,还可促进 5- HT 递质的功能改善,从而通过改善神经运动功能来促进脑损伤后的功能再塑.我们采用氟西汀灌胃发现:治疗后的形觉剥夺弱视猫潜伏期逐渐缩短,波幅逐渐增高,以氟西汀治疗12 周后 P 波潜伏期最短,成比组有统计学意义.MaYa 课题组[6]在 4 周内喂给一些模拟人类弱视的老鼠每日剂量的氟西汀,成年老鼠的视觉获得部分恢复.研究发现经治疗的鼠的视觉中枢活动活跃,同时还显示它们的表现和视力正常的老鼠一样,似乎表明氟西汀可以使成年大鼠脑中与视觉有关的神经元回复到它们年轻时的更具可塑性的状态,并且即使大脑处于成年期,也能使其从某些类型的视觉损害中康复.
有关作用机理的推测:认为氟西汀通过改善大脑神经元可塑性状态而发挥疗效,这使得视知觉系统发育自身眼-脑间的合理联系,能够增加大脑神经细胞的可塑性理论的研究-即这些大脑神经细胞能够建立或打破他们之间的突触联系,这就像是在准确视力的基础下进行的神经线路连接过程.
该项发现提示大脑可塑性是这个过程的关键部分,有助于治疗弱视.以往的研究一直认为视皮层可塑性只存在于幼年动物视觉发育关键期内,但近几年来的研究使我们逐渐认识到成年动物视皮层仍然存在着一定程度的可塑性,当外界视觉环境发生改变时,这种可塑性可能会被再激活并表现出来,这样的视皮层可塑性在动物实验和人类的医疗实践中都有表现[7- 9].
通过该实验发现,氟西汀治疗形觉剥夺弱视模型猫能使其视觉诱发电位 P 波潜伏期缩短,波幅增加,并具有统计学意义,其变化过程类同于人类弱视儿童P- VEP 的 P 波的潜伏期和波幅的变化趋势.下一步我们将探索氟西汀的最佳浓度,期望能为临床治疗弱视提供新的治疗手段.
参 考 文 献
1. 高建华,张东果 ,郭守一,等.视觉剥夺对幼猫图形视觉诱发电位的影响.眼视光学杂志,2002,4(4):233.
2. Chen JY, Hood DC, Odel JG, et al.The effect of retinalabnormalities on the multifocal visual evoked potential.Inves-tigative ophthalmology & visual science,2006,47(10):4378.
3. Brendan T, Barrettt, Arthur Bradley.Understanding theNeural Basis of Amblyopia. Neuroscientist,2004,10:106.
4. 殷小龙,毛欢文,鲁纯葆,等 . 弱视儿童图形视觉诱发电位的分析 . 江西医学院学报,2002,42(6):158.
5. Trachtenberg JT,Trepel C. Rapid extragrenular plasticity inthe absence of thalamocortical plasticity in the developingprimary visusl cortex . Science,2000,287(5460):2029.
6.MaYa Vetencourt JF,Viegi A et al. The antidepressant fluox-etine restores plasticity in the adult visual Cortex.Science,2008,320(5874):385.
7. Kaas JH. Plasticity of sensory and motor maps in adult mam-mals. Annual review of neuroscience,1991,14:137.
8. Gilbert CD. Adult conical dynamics.Physical review,1998,78(2):467.
9. Tagawa Y,Kanold PO,Majdan M,et a1. Multiple periodsof functional ocular dominance plasticity in mouse Visualcortex.Nature Neuroscience,2005,8(3):380.





