断奶仔兔的生长发育尚未完全,抵抗外界环境的调节机能较差,容易产生腹泻、腹胀和球虫等疾病,降低了日增重和饲料报酬,发病率和死亡率增加. 饲养者往往通过添加抗生素来增强仔兔的抗病力,而抗生素在预防和治疗家兔疾病上的滥用现象,常常引起细菌耐药性的产生和肠道菌群紊乱,造成二次感染. 微生态制剂以其无污染、无残留、安全可靠、不产生抗药性等优点广泛应用于畜牧业. 研究者发现,饲喂微生态制剂可以调节仔鹅盲肠微生态菌群,提高机体免疫力和生长性能,同时增强断奶仔兔的抗病力,降低料肉比,减少腹泻率,提高断奶仔兔的生产性能.
1997 年变性梯度凝胶电泳技术 (DGGE)首 次应用于微生物生态学研究, 并证实该技术在研究自然界微生物群落遗传多样性和种群差异方面具有显着优越性. 在畜牧业养殖中,DGGE 逐渐应用于牛、羊、猪和鸡等动物肠道菌群多态性和系统发育关系的研究,在兔肠道的研究中应用还较少.
白秀娟等(2012)研究结果表明,PCR-DGGE 完全可以用于断奶后仔兔肠道菌群结构的研究, 从而弥补了传统微生物分离培养方法费时费力和精确度差的缺陷. 本文旨在通过测定添加微生态制剂后回肠 SIgA 的分泌量, 探讨微生态制剂对断奶仔兔肠道免疫功能的作用, 并从分子水平探讨肠道菌群的变化,为其在畜牧业中应用提供参考.
1 材料与方法
1.1 材料
肽菌素:购自山东宝来利来生物工程股份有限公司,主要由高活性抑菌型枯草芽孢杆菌、 乳酸菌及其代谢产物抗菌肽组成,活菌总数≥1.5×108cfu/g.
常康泰:购自山东宝来利来生物工程股份有限公司,主要由高效抑菌型芽孢杆菌、乳酸菌、细菌素、抗菌肽及肠道修复因子组成,活菌总数≥3.0×108cfu/g.
布恩氏固定液、苏木精和伊红染色液:由山东宝来利来生物工程股份有限公司生物研究院实验室自制.
SIgA 检测试剂盒:购自普天试剂公司.
粪便基因组提取试剂、PCR 扩增试剂: 购自 TIANGEN公司.
实验设备: 切片机、 酶标仪、PCR 仪 、DGGE 电泳仪(Bio-Rad)、脱色摇床.
1.2 试验设计与日粮组成
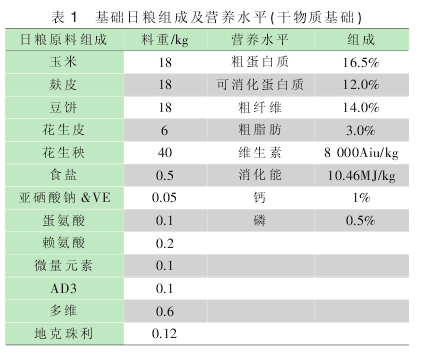
试验采用单因子完全随机区组设计, 选用刚断奶仔兔120 只,平均体重为 513g. 随机分为 4 组,每组 3 个重复,每个重复 10 只兔. 对照组饲喂基础日粮 (参考正常兔料营养标准), 试验 1~3 组分别在基础日粮的基础上添加肽菌素(120 g/t)、复合肽菌素 (120 g/t)和 锌酵母 (150 g/t)以 及常康泰(500 g/t). 基础日粮根据当地肉兔的营养水平配制,日粮组成及营养水平见表 1.
1.3 饲养管理
断奶仔兔采用笼养方式,每笼 2~3 只. 试验期间仅提供颗粒料,不饲喂其他精青粗饲料. 每天早晚定时喂料,自由采食、饮水. 搞好兔舍卫生工作,保证兔舍温暖、干燥、清洁,统一免疫和驱虫.
1.4 样品采集
试验期结束后,将各组兔处死后解剖,取一段回肠,冲洗干净后于 10%的福尔马林溶液中固定用于观察小肠绒毛,另取一段回肠(连同内容物)-80 ℃保存,用于测定 SIgA 的表达量,最后取盲肠内容物,将每组每个重复内 10 只兔的盲肠粪便做等份混合,即每个组有 3 个盲肠粪便样品,再将这3 个样品做等份混合形成第 4 个样品,即每组有 4 个样品用于检测微生物菌群的变化.
1.5 测定指标及方法
1.5.1 兔回肠绒毛观察
截取一小段兔回肠包埋在石蜡中,使用切片机横向切片后脱蜡,布恩氏固定液固定 5 min 后乙醇洗涤,苏木精、伊红染色各 30 min,最后用加拿大树胶封片. 置 45 ℃温箱中烘烤过夜后进行镜检.
1.5.2 兔回肠 SIgA 抗体水平的测定
准确截取 1 cm 肠段,用剪刀纵向剖开. 加入 5 mL PBS缓冲液,漩涡震荡 1 min. 8 000 r/min 离心 5 min,取上清液,移入新的离心管. 取的上清液 12 000 r/min 离心 10 min 后取上清液为待测样品,用 ELISA 试剂盒检测.
1.5.3 兔肠道菌群结构的变化
每个试验组称取粪便样本 180~220 mg 至干净的 2 mL离心管,离心管置于冰盒上. 按照粪便基因组提取试剂盒说明书的步骤提取兔粪便总 DNA. 以提取的 DNA 直接作为PCR 模板,采用细菌 16S rRNA 基因 V3 区通用引物对 :GC-357F (5'-C GCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGG CCTACG GGAGGCAGCAG-3') 和 517R (5'-ATTACCGCGGCTGCTGG- 3'), 引物由上海生工合成. PCR反应体系 25 μL:2×Taq mix 12.5 μL,上、下游引物(10 umol/L)各 1 μL,模板 DNA1 μL,无菌双蒸水 9.5 μL. PCR 反应程序为: 94 ℃预变性 4 min;94 ℃变性 1 min,65 ℃退火 45 s,退火温度每个循环降低 0.5 ℃,当退火温度降至 55 ℃后,再以该温度扩增 15 个循环;最后 72 ℃延伸 8 min. PCR 产物用 1%的琼脂糖凝胶(EB 染色)紫外下进行检测.
配制变性梯度为 30%~70%的 DGGE 胶,电泳时加入 10μL 的样品和少许 loadingbuffer,打开电泳仪,设置电压为 80V,时间为 668 min. 电泳结束后,进行硝酸银染色,显影. 相机 拍 照 后 采 用 FastStone Capture 软 件 进 行 截 图 , 使 用Quantity One 软件(Bio-Rad)对 DGGE 指纹图谱进行丰富度分析和聚类分析.
用无菌手术刀将 DGGE 图谱中部分优势明显、 条带清晰且亮度高的特异性条带从凝胶上切下, 加入 40 μL 去离子无菌水, 将胶块捣碎,4 ℃过夜. 取液体做模板进行 PCR扩增,引物为去掉"GC 夹"的 GC-357F 和 517R,PCR 体系和条件同上. 扩增产物大小在 161-201bp,送上海铂尚生物技术有限公司测序,得到的测序结果和 NCBI 中 GenBank 数据库中 BLAST 比对分析.
1.5.4 数据处理
数据先使用 Microsoft Excel 整理,再通过统计软件 SPSS11.5.0 进行单因素方差分析 (One-way Anova),用 LSD 法 比较分析差异, 显着标准为 P<0.05, 结果以直方图或者平均数±标准误差表示.
2 结果与分析
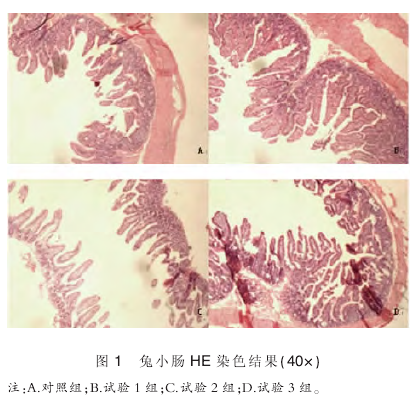
2.1 兔回肠 HE 染色结果
由图 1 可知,相对于对照组,三个试验组的小肠绒毛高度明显上升,隐窝变深,吸收营养物质的有效面积明显增大.
从形态上看,对照组小肠绒毛形态呈现叶状,试验 1 组小肠绒毛为舌状,试验 2 组和 3 组的小肠绒毛形态为指状.
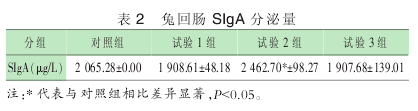
2.2 兔回肠 SIgA 检测结果
由表 2 可以看出, 与对照组比, 试验 1 组、3 组兔回肠SIgA 表达无显着变化, 而试验 2 组 SIgA 表达量显着增加 .
说明试验 2 组微生态制剂的添加可以提高断奶仔兔的免疫力,增强机体的抗病能力.
2.3 兔肠道菌群结构的变化
2.3.1 PCR 扩增结果
16srDNA 的扩增产物经琼脂糖电泳后结果如图 2 所示,每个泳道只有 1 条亮带,未产生非特异性条带,基因扩增片段大小均在 161~201bp, 参考 DNA marker 可看出各目的基因的片断大小与设计相符, 表明所扩增片段为特异性目的片段.
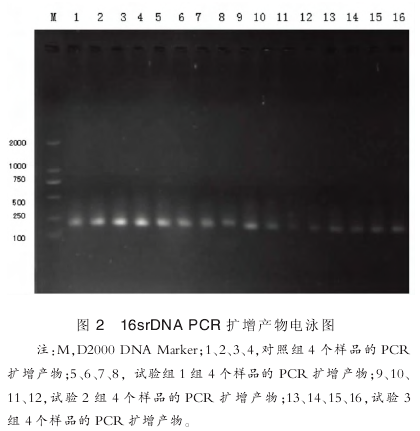
2.3.2 DGGE 指纹图谱分析
各样品 DGGE 图谱如图 3 所示. 每一泳道代表一个样品,每个泳道内的条带代表一种细菌,条带的亮度代表该菌的相对丰度. 丰富度(S)为 DGGE 图谱中每一泳道的条带数. 如表 3 所示,盲肠内的菌种数接近 30 种,而相对于对照组,试验各组无明显差异性.
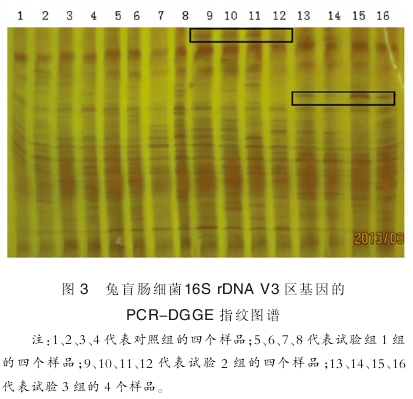
2.3.3 聚类分析
由图 4 可以看出 1、2、3、4 的菌群相似性均在 0.53 以上, 其中 2、3 样品的相似性为 0.81;5、6、7 的相似性在 0.83以上;9、10、11、12 样品的相似性在 0.58 以上;13、14、15 的相似性也在 0.48 以上, 说明组内各样品之间的微生物菌落差异不大,动物建模较为成功. 同时,可以看出对照组和试验一组的样品之间相似性在 0.53 以上,对照组、试验 1 组、2组与试验 3 组的相似性也在 0.48 以上, 说明这三个组微生物菌群变化不大;而对照组、试验 1 组、试验 3 组与试验 2 组的相似性则只有 0.39,说明试验 2 组的处理导致了仔兔肠道菌群的明显变化.
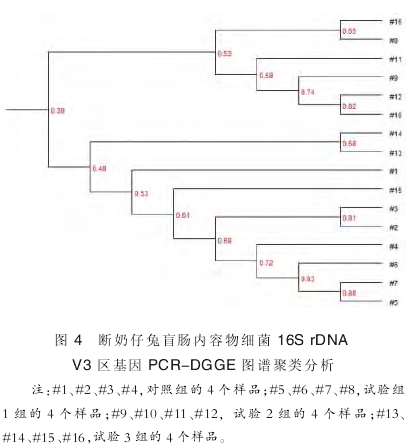
2.3.4 16S rDNA 序列测序结果
图 2 黑框内的条带分别为试验 2 组和试验 3 组中亮度较高的差异条带,选取该条带割胶测序. 测序结果表明,试验 2 组差异菌种与 Bifidobacterium animalis 亲缘关系最近,相似性为 99%,为动物双歧杆菌菌属;试验 3 组差异菌种与Lactobacillus plantarum,相似性为 99%,为植物乳酸菌菌属.
3 讨论
DGGE 是由 Fisher 等发明用于检测 DNA 突变的一种电泳系统, 它是利用序列不同的 DNA 片段在聚丙烯酰胺凝胶中解链温度不同的原理, 通过梯度变性胶将 DNA 分开(FISCHER S 等,1983). 与其它电泳系统相比,它不是基于核酸分子量的不同将 DNA 片段分开, 而是根据序列的不同,将片段大小相同的 DNA 序列分开. 理论上认为,只要电泳条件(如电泳时间、变性剂梯度、电压等)精确控制,DNA 片 段可分辨到一个碱基的差异 . 利用 DGGE 指 纹图谱分析微生物菌群,具有快速精确、无需培养、检测度高的优势.
本研究结果说明,微生态制剂可以促进仔兔小肠绒毛高度的上升和隐窝的加深,扩大营养物质吸收面积,提高饲料报酬. 以往研究也发现,在断奶应激情况下,饲料中添加微生态制剂可提高仔兔的生产性能和免疫功能, 维护断奶仔兔肠道菌群平衡和促进肠道发育, 这与本次试验结论相一致. SIgA 存在于不同组织的粘膜上,在结构上比 IgA 多一条氨基链,由局部粘膜自行合成,不受血清中抗体调节,回肠中含有大量的浆细胞可以分泌 SIgA, 它可以与抗原发生结合反应,从而阻止肠道粘膜外来微生物的定植,抵抗病原菌的感染. 锌酵母和肽菌素的联合使用可以提高兔回肠 SIgA 水平,增强兔肠道对病菌的抵抗力. 丰富度分析说明兔盲肠内微生物种群丰富,菌种接近 30 种,而添加微生态制剂后不会引起盲肠内微生物群落的急剧改变,在一定程度上有效地保持了兔盲肠内的微生态平衡. 以往研究发现,猪和鸡不同个体的肠道内容物菌群结构有很大差异,研究者常常将同一处理组的样本进行混合来减少平行样本间的差异性. 为减少组内样本之间的差异性,该实验将每组的 3 个样本进行了等量混合,形成第 4 个样本,聚类分析说明组内样本差异性较小,保证了数据的可用性. 结果显示,基础日粮中单独添加肽菌素或者添加常康泰不能有效地改变兔盲肠内的菌群结构,而将锌酵母和肽菌素联合组方后,兔回肠 SIgA 水平显着上升,盲肠内的菌群结构发生了较大的变化. 锌是兔生长所必需的一种微量元素,是体内多种酶的重要组成成分. 梅绍锋等(2009)指出锌的添加能提高胃蛋白酶和胰淀粉酶活性,降低盲肠大肠杆菌、 乳酸杆菌和双歧杆菌数量; 张之申等(1992)发现锌酵母的添加可以促进锌在家兔体内的吸收,提高断奶仔兔的免疫力和成活率. 因此,可以推断锌酵母的添加可以协同肽菌素提高断奶仔兔的肠道免疫水平,改善肠道菌群结构.
测序结果证明,基础日粮中添加锌酵母和肽菌素组方或者添加常康泰分别增加了乳双歧杆菌和植物乳酸菌的比例,张振瑞等(2007)通过鉴定健康家兔肠道内的益生菌,发现肠道内优势菌群包括梭杆菌属、双歧杆菌属、乳杆菌属、明串球菌属、拟杆菌属,与本次研究结果相一致. 因此,基础日粮中添加锌酵母和肽菌素组方或者常康泰可以促进断奶仔兔小肠的粘膜免疫,提高盲肠内有益菌群的优势度,从而有效地抑制有害菌的发生和繁殖.
参考文献:
[1] 赵泮峰,任战军,王洪阳. 早期断乳幼兔腹泻内因分析[J]. 饲料工业,2010,31(23):52-54[2] PAKANDL M. Coccidia of rabbit: a review [J]. FOLIA PARASITOLOGICA,2009,56(3): 153-166.
[3] 董巍,张玉杰,林丰海,等. 微生态制剂对黑龙江仔鹅盲肠菌群变化的影响研究[J]. 中国畜牧兽医,2007,34(8):20-22.
[4] 李振. 微生态制剂对断奶仔兔生产性能及免疫的影响[J]. 中国微生态学杂志,2009,21(8):718-720.
[5] GIDERME T,JEHL N,SEGURA M,et al. Microbial activity in the caecum of the rabbit around weaning: impact of a dietary fiberdeficiency and of intake level [J]. Animal Feed Science and Technology,2002,99: 107-118.
[6] MYER R,FISCHER S,LERMA L,et al. Nearly all single base substitutions in DNA fragments joined to a GC-clamp can be detectedby denaturing gradient gel electrophoresis [J]. Nucleic Acids Res,1985,13(9):3131-3145.
[7] ZOETENDAL E,COLLIER C,KOLKE S,et al. Molecular ecological analysis of the gastrointestinal microbiota: A Review [J]. J Nutr,2004,134: 465-472.
[8] 白秀娟,刘诚刚,杜智恒,等. PCR-DGGE 技术分析断奶仔兔肠道微生物菌群结构及多样性[J]. 东北农业大学学报,2012,43(9):64-69.
[9] LAKNER V,BROCK J. Secretory immunoglobulin A [J]. Allerg Immunol(Leipz),1977,23(3): 147-58.
[10] GONG J,SI W,FORSTER R,et al. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny inthe chicken gastrointestinal tracts: From crops to ceca [J]. FEMS Microbiol Ecol,2007,59: 147-157.
[11] LI M,GONG J,COTRILL M,et al. Evaluation of QIAamp DNA mini stool kit for microbial ecological studies [J]. J MicrobiolMethods,2003,54: 13-20.
[12] SIMPSON JM,MCCRACKEN V,GASKINS H,et al. Denaturing gradient gel electrophoresis analysis of 16S ribosomal DNAamplicons to monitor changes in fecal bacterial populations of weaning pigs after introduction of Lactobacillus reuteristrainMM53 [J]. Appl Environ Microbiol,2000,66: 4705-4714.
[13] 白国勇. 乳酸锌的研究进展及在兔营养中的应用[J]. 四川畜牧兽医,2007,8:34-35.
[14] 梅绍锋,余冰,鞠翠芳,等. 高锌和高铜对断奶仔猪生产性能、消化生理和盲肠微生物数量的影响[J]. 动物营养学报,2009,6.
[15] 张之申,石玉诚,郭锡勇,等. 富锌酵母在兔体内的生物利用度研究[J]. 中国医药工业杂志,1992,23(8):358-359.
[16] 张振瑞,张 力,陈桂银,等. 健康家兔肠道益生菌的分离鉴定[J]. 中国养兔,2007,2:21-24.





