����ժ Ҫ����Ŀ�� ̽���������鵼�������ǵ�����������������2 (Slit2) ����������תȾ�Թ������ʸ�ϸ�� (BMSCs) ����ϸ��ת�������á����� ���û�������\��, ����Slit2����������;������תȾBMSCs;������ʽϸ���Ǽ��BMSCs�ı��浰��, RT-PCR��Western-blot�������հ����顢�����岡������������岡������ϸ�����浰������ά���Ե��� (GFAP) ����Ԫ������ϩ����ø (NSE) �ı����� ��2��BMSCs����CD44��CD29��CD34��CD45�ı����ʷֱ�Ϊ60.2%��58.3%��3.4%��2.6%, ������תȾЧ��Ϊ90%���ϡ��հ����顢�����岡������������岡�����GFAP��NSE mRNA�͵�����Ա������Ƚ�, ������ͳ��ѧ���� (F=9.089��13.893, P<0.05) ;���й��������岡�����GFAP��NSE mRNA�͵�����Ա��������Ը��ڿ����岡���� (P<0.05) , �����岡����Ϳհ�����Ƚϲ����������� (P>0.05) ������ Slit2�����������������BMSCs����ϸ��ת����Ч��, Ϊ�����˺ͼ��������ṩ����ϸ����
�����ؼ��ʣ�������������; ����; ����ʻ���ϸ��; ����ԭ��ά���Ե�����; �����ͪ��ˮ��ø;
����Abstract����Objective To explore the effect of lentivirus-mediated overexpression of Slit2 on the transformation of bone marrow mesenchymal stem cells (BMSCs) into neural cells. Methods A Slit2 overexpression vector was constructed and used to transfect BMSCs via lentivirus.Flow cytometry was used to detect surface proteins of BMSCs.RT-PCR and Western blot were used to determine the expression of glial fibrillary acidic protein (GFAP) and neuron-specific enolase (NSE) on the surface of neural cells in blank control group, empty lentiviral vector group, and overexpression lentiviral vector group. Results The expression of CD44, CD29, CD34, and CD45 was 60.2%, 58.3%, 3.4%, and 2.6%, respectively, on the surface of the second generation BMSCs.The transfection efficiency of lentivirus was over 90%.There were significant differences in relative mRNA and protein expression of GFAP and NSE between the blank control group, the empty lentiviral vector group, and the overexpression lentiviral vector group (F=9.089-13.893, P<0.05) .The overexpression lentiviral vector group had significantly higher relative mRNA and protein expression of GFAP and NSE than the blank control group and the empty lentiviral vector group (P<0.05) , while there were no significant differences between the blank control group and the empty lentiviral vector group (P>0.05) . Conclusion The Slit2 overexpression plasmid can promote the efficiency of the transformation of BMSCs into neural cells, which provide seed cells for nerve injury and spinal cord injury.
����Keyword����nerve growth factor; bone marrow; mesenchymal stromal cells; glial fibrillary acidic protein; phosphopyruvate hydratase;
�����������ʸ�ϸ�� (BMSCs) ���ж���ֻ�DZ��, ���ض�������, ���Էֻ��������ڵĸ���ϸ��, ��������ϸ������ϸ���Լ���ϸ����[1,2,3,4]���о������ʾ, BMSCs���ض��յ������¿���ת������ϸ��, �����ܴٽ������˵���;����BMSCs��Դ�ڲ�������, ��ֲ����Ա����ųⷴӦ[5];����, BMSCs�ܷ��ڶ���ϸ������, ��Ϊ�����ṩ��һ���µ�˼·[6]���������ǵ�����������������2 (Slit2) ��������ϵͳ���������е�һ����ͻ������������, �ɿ�������ͻ��֧�γ��Լ���ϸ��Ǩ��, �ܴٽ�BMSCs��о���Ԫ�ֻ�, ��������������������[7,8,9]��������, �ڽ�ͨ�¹��з����������˺ͼ������˱Ƚϳ���, ����������˵�����һֱ���ٴ��ϵ��ѵ㡣���о���Slit2�������������������鵼��BMSCs��, ̽���������鵼Slit2������תȾ�ٽ�BMSCs����ϸ��ת��������, �Ӷ�Ϊ�ٴ������˲��������ṩ�µ�˼·��
����1�� �����뷽��
����1.1�� ʵ�����
����̥ţѪ�幺��Gibco��˾;DMEM��2.5g/L��øTrypsin��������Invitrogen��˾;Trizol��������Invitrogen��˾;�ȷ¡��Ҵ�����ת¼�Լ��й�������ABI Applied Biosystems��˾;Real-time PCR�ǹ�������bio-rad��˾��
����1.2�� ʵ�鷽��
����1.2.1�� ��BMSCs�ķ��������
����ѡȡ20������־Ը��, ��8��, Ů12��, ����Ϊ20��38��, ƽ�� (27.48±7.83) �ꡣʵ�鷽������˱�Ժ����ίԱ�����, ����־Ը�߾�ǩ��֪��ͬ���顣־Ը����ǰ����, �ų����ס��Բ������̲��ȴ�Ⱦ�Լ���;����ȡ����λ, ��¶˫���ĺ��ϼ�, ����, �̽�, �������20mL�ֲ�����, ���ùǴ�����ɼ�־Ը�ߵ����ʹ���, ����1��3����ȼ��뺬0.01mol/L̥ţѪ�塢100kU/L��ù�ص�DMEMϸ������Һ, ��ֻ��Ⱥ�, ����25cm2��Hank����ƿ��, ���ò�������������, 3d��ȥ������ƿ�е���֬������, ֮��ÿ3d��1��ϸ������Һ��BMSCsϸ���ں��ʴ�70%��80%����д���������
����1.2.2�� BMSCs�ļ���
����������ʽϸ���Ǽ���2��BMSCs���浰��CD29��CD44��CD71��CD90��CD106�ı���;���õ��ù�ѧ�����۲�BMSCs��ϸ����̬��
����1.2.3�� ������Slit2���������幹��
����������Slit2�������������Ϻ���������ѧ��������˾�ϳ�, ѡ��pHBLV-U6-ZsGreen-PGK-Puro��Ϊ������¡���塣������Slit2�����������¡�����ΪXhoI/BamHI, ���ΪENST00000005893��
����1.2.4�� ������ϸ��תȾ��ʵ�����
����ȡ��3��BMSCs����ʵ�顣��ϸ�����ֵ�6�װ�, ��ϸ���ں���Ϊ50%ʱ����������תȾ, ����תȾ˵����, MOI=50, �������ζ�Ϊ3×107����BMSCsϸ���ֳ�3�顣�հ�����:δתȾ������;�հ�������������:תȾ����Slit2����������������;Slit2������������������:תȾSlit2������������������1.2.5ʵʱӫ�ⶨ��PCR (RFPCR) �������ԭ��ά���Ե����� (GFAP) ����Ԫ������ϩ����ø (NSE) mRNA����3��ϸ���ں��ʴ�80%, ��2.5g/L�ȵ���ø���������15mL���Ĺ���, ����1mL��Trizol RNA iso (Takara, �ձ�) , ����Trizol��ȡ��, ��ȡ��RNA��������ת¼�Լ�����ת¼Ŀ��RNA, ���õ���cDNA������-80������С�����ӫ�ⶨ��PCR�Լ���˵����Ҫ��, ��Ӧ��ϵΪ25μL, �ں�:SYBR Premix Ex TaqTM�� (2×) 12.5μL, PCR Forward Primer (10μmol/L) 1.0μL, PCR Reverse Primer (10μmol/L) 1.0μL, cDNAģ��2.0μL, ddH2O 8.5μL��Ӧ��FS 2000ϵͳPCR������25μL�ķ�Ӧ��ϵ���з���, ����������PCR��Ӧ����, ��Ӧʱ��ѡ��120 min;����Ԥ����:95�桢30s;Circle 1:95�桢30s;Circle 2:60�桢30s;�ܽ�����:95�桢5s, 60��1 min;����:50�桢30s����¼��������CTֵ���ڲλ����������Ϻ�������˾�ϳɡ�����1����2-����CT������GAPDH��GFAP��NSE����Ա�������
����1.2.5��ʵʱӫ�ⶨ��PCR (RFPCR) �������ԭ��ά���Ե����� (GFAP) ����Ԫ������ϩ����ø (NSE) mRNA����
����3��ϸ���ں��ʴ�80%, ��2.5g/L�ȵ���ø���������15mL���Ĺ���, ����1mL��Trizol RNA iso (Takara, �ձ�) , ����Trizol��ȡ��, ��ȡ��RNA��������ת¼�Լ�����ת¼Ŀ��RNA, ���õ���cDNA������-80������С�����ӫ�ⶨ��PCR�Լ���˵����Ҫ��, ��Ӧ��ϵΪ25μL, �ں�:SYBR Premix Ex TaqTM�� (2×) 12.5μL, PCR Forward Primer (10μmol/L) 1.0μL, PCR Reverse Primer (10μmol/L) 1.0μL, cDNAģ��2.0μL, ddH2O 8.5μL��Ӧ��FS 2000ϵͳPCR������25μL�ķ�Ӧ��ϵ���з���, ����������PCR��Ӧ����, ��Ӧʱ��ѡ��120 min;����Ԥ����:95�桢30s;Circle 1:95�桢30s;Circle 2:60�桢30s;�ܽ�����:95�桢5s, 60��1 min;����:50�桢30s����¼��������CTֵ���ڲλ����������Ϻ�������˾�ϳɡ�����1����2-����CT������GAPDH��GFAP��NSE����Ա�������
������1 Ŀ�Ļ������������

����1.2.6�� Western-blot���GFAP��NSE����Ե��ױ���ˮƽ
����3��ϸ��������, �ֱ����RIPAϸ���ѽ�Һ1mL, �ڱ����ѽ�30min, �ռ���1.5mL�����Ĺ���, ��4��Ԥ������Ļ������� (5 000r/min, 5min) , ��ȡ����, ��95����к�, �ռ����ױ��á��Ʊ�Ũ����10mL, ���뽺20mL���������Ӿ����, תĤ����4������бܹ������ҹ (>12h) ��GFAP��NSEС����һ�� (Abcam��˾) 1��10 000ϡ�ͺ������������Ĥ;��2����PBSTϡ��ҺϴĤ, �ظ�3��, ��ɽ��С��IgG���� (�����칫˾) 1��1 000ϡ�ͺ����1.5h, ʹ��PBSTϡ��ҺϴĤ, �ظ�3��, Ȼ����DAB��ӰҺ������Ӱ��Ӧ��Image J������������������
����1.3 ��ͳ��ѧ����
����Ӧ��SPSS 16.0��������ͳ��ѧ����, �������Ͻ������ʽ��ʾ���������ݼ�Ƚϲ��õ����ط������, ��������Ƚϲ���q���顣
����2 �����
����2.1�� ��BMSCs�������ͼ���
��������48h��, ԭ��BMSCs��ʼ����, ϸ�������λ��߶����;��������2��ϸ��������״���߷���״����, ϸ����ֳѸ�١���ʽϸ���Ǽ����ʾ, ��2��BMSCs���浰��CD44��CD29�����ʷֱ�Ϊ60.2%��58.3%, CD34��CD45�����ʷֱ�Ϊ3.4%��2.6%��
����2.2�� �������鵼Slit2����������תȾBMSCs
���������ʾ, ��������תȾЧ�ʽϸ�, Ϊ90%����, ���������̶��Ϸ���Slit2�Ĺ��������á�
����2.3�� ����GFAP��NSE mRNA�͵��ױ���Ƚ�
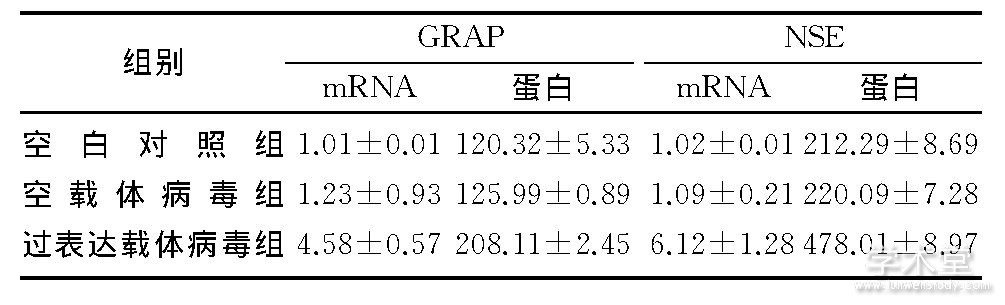
�����հ����顢�����岡������������岡�����GRAP��NSE mRNAˮƽ�͵��ױ������Ƚϲ�����ͳ��ѧ���� (F=9.089��13.893, P<0.05) , ���п����岡����Ϳհ�����Ƚϲ����������� (P>0.05) ;���������岡����Ϳ����岡������Ƚ�, �������ͳ��ѧ���� (q=2.001��3.897, P<0.05) ������2��
������2 ����GRAP��NSE mRNA�͵��ױ���
����3 ������
����������, ���Ž�ͨ�¹ʵĶ, �ڽ�ͨ�¹��з����������˺ͼ������˱Ƚϳ���[10,11,12], ����������˵�����һֱ���ٴ��ϵ��ѵ�[13]��BMSCs���ж�ֻ�DZ��, ������о�����, BMSCs���Էֻ�����ϸ��[14,15,16,17,18]������������Slit2�Ĵ̼���, BMSCs��ת������ϸ��, ����Ч�ʲ���[19,20]�����о����û�������\��, ����Slit2�Ĺ���������, �����ø�תȾЧ�ʵ�������תȾBMSCs, ̽��BMSCs����ϸ����Ч��ת����ʵ�鷽��, Ϊ�ٴ��������˺ͼ��������ṩ����ϸ����
����Slit2������������������ϸ����������������[21]�����о��������, Slit2ͨ��Ũ���ݶ����鵼��ϸ����Ǩ�ƺ�����, ����ϸ�����������ų��Ե�������, ͬʱ���ɴٽ��о���ϸ��������ά��֧�γ�[22,23,24]����ʾ���������ӿ��Դٽ���Ԫ��ϵϸ���ķֻ�[25]�����о�Ӧ�û�������\������Slit2����������, Ӧ��������תȾBMSCs, �����ʾ, ��������תȾЧ�ʽϸ�, Ϊ90%����, �������̶ȷ���Slit2�Ĺ��������á�
����GFAP��NSE������ϸ���ı�ʶ����, ����GFAP�����ν���ϸ����ı�־��, ��Ҫ�ֲ���������ϵͳ�����ν���ϸ��, ����ϸ���ǼܵĹ��ɲ�ά��������ǿ��[26,27]��Ѫ��NSE����Ԫ�����ڷ���ϸ�������е�һ�����Ե���ø[28]�����о�ѡ������������ϸ���������Ա�־��, ����RT-PCR��Western-blot��ʵ�鷽��, �ӻ���͵������������ϼ��հ����顢�����岡������������岡�����GRAP��NSE�ı���ˮƽ, �����ʾ, ���������岡����GRAP��NSE����͵��ı���ˮƽ�����Ը��ڿ����岡����Ϳհ�����, ������ͳ��ѧ���塣�Ӷ�����֤���ӻ���ˮƽ�Ͻ�Slit2�Ĺ���������ͨ���������鵼�ķ���תȾBMSCs, �������BMSCs����ϸ����ת��Ч��, �ʶ���ϸ���ı����GFAP��NES�ı������������ߡ�
������������, Slit2�����������������BMSCs����ϸ��ת����Ч��, Ϊ�ٴ��������˺ͼ������������ṩ����ϸ����
���������
����[1]���, ͯ�ཨ.������Ѫ����ҩѪ���Ԥ������������������ʸ�ϸ�������Ƿֻ���������Ѫ�����Ϲ������ʸ�ϸ�����ƴ���ϥ�ǹؽ���ʵ���о�[J].��ҽ����, 2018, 30 (1) :6-11.
����[2]������, ��÷, ����, ��.TNF-α��С��������ʸ�ϸ��������������Ӱ��[J].�ൺ��ѧҽѧԺѧ��, 2015, 51 (2) :169-171, 174.
����[3]�Է�Ӣ, ������, ��梷�.Wnt3a���˹������ʸ�ϸ���ɹǷֻ���Ӱ�켰�����о�[J].�й�ҽ�ƴ�ѧѧ��, 2018, 47 (7) :617-621.
����[4]¬־ΰ, ����, ����Ӣ, ��.���ζ��ǶԵ������������ʸ�ϸ�����ֻ�DZ�ܵ�Ӱ��[J].�й�ҩ��ѧͨ��, 2017, 33 (7) :950-955.
����[5]�ż���, ����Ԫ, ����, ��.�����յ�����������ʸ�ϸ������ϸ���ֻ����о���չ[J].����ҽ�����Ѫ�ܲ�������־, 2017, 5 (27) :32-33.
����[6]���, ��ˮ��, ����, ��.�������ʸ�ϸ�����Ƹ������Զ������ʵ�鼼����ϵ�����Ҫ[J].�л�����ѧ��־, 2017, 15 (3) :350-356.
����[7]������, ������, ������, ��.ȱ����ȱѪ�յ������������ʸ�ϸ������Slit2�����Ӱ��[J].����ҽѧ�������ٴ�, 2017, 21 (2) :189-194.
����[8]����, �Ž���, ��Դ, ��.��Ǩ�Ƶ���Slit2��������ʸ�ϸ����Ѫ�������Ĺ�ϵ[J].�й���֯�����о�, 2014, 18 (37) :6034-6039.
����[9]��Դ, ����, ������, ��.��������Slit2���ι������ʸ�ϸ���ı����о�[J].�з���������־, 2015, 32 (12) :1084-1088.
����[10]���, �ź���, �˱�, ��.��̴ٽ��������˺����ָܻ��Ļ��Ƽ�����ź�ͨ·������[J].�й�����������ʵ��, 2017, 23 (6) :641-644.
����[11]������, ��ѩ��, ����, ��.�м��������ϼ�ǿ�������Դ����Լ������˺���ϸ�����������ʵ���о�[J].��ҽҩ��Ϣ, 2017, 34 (1) :92-95.
����[12]����ϼ, ����, ��˧, ��.��Ƶ��̼���Ͽ���ѵ�����Ƽ���������Դ��������Ч�۲�[J].�й��ٴ��о�, 2017, 30 (1) :116-119.
����[13]����, ������, ������, ��.���ᰱ�������ϴ������ǿ������������ƴٽ��������ۺϲ����������������ָܻ����õij���̽��[J].�й��ٴ�ҩ��ѧ������ѧ, 2017, 22 (8) :937-942.
����[14]�����, ����, �Ծ���, ��.�������ʸ�ϸ���Ը���������ϸ�����˵�������[J].��ž�ҽѧ��־, 2017, 42 (5) :377-382.
����[15] ¬־ΰ, ����, ����Ӣ, ��.���ζ��ǶԵ������������ʸ�ϸ�����ֻ�DZ�ܵ�Ӱ��[J].�й�ҩ��ѧͨ��, 2017, 33 (7) :950-955.
����[16]�Ź���, ���ӄ�, ����, ��.�Ȼ��ͨ����������ͨ·�ٽ�����������ʸ�ϸ������ϸ���ֻ�[J].�й�����������־, 2017, 33 (12) :2128-2133.
����[17]�ϼѼ�, ����, �, ��.��ϸ��𤸽���Ӷ�С��������ʸ�ϸ��𤸽��Ǩ�Ƽ�ϸ����̬��Ӱ��[J].����ҽѧ���ٴ�, 2017, 37 (8) :1082-1087.
����[18]����, ����, ¬��.��ѧ���������յ�����������ʸ�ϸ��������ϸ���ֻ�[J].���\��ͨѶ, 2017, 28 (6) :793-797.
����[19]���Ǿ�, ����.�������ʸ�ϸ���������е��о���չ[J].�������빦���ؽ�, 2016, 11 (5) :426-428.
����[20]����, ������, ������, ��.Let-7c����������Թ������ʸ�ϸ�������յ��ֻ�Ϊ��ϸ����Ӱ��[J].�й���֯�����о�, 2016, 20 (1) :20-25.
����[21]��ʿ��, �����, �ſ���, ��.BALB/cС��SLIT2��ROBO4������֯�����Ա�����ֲ����о�[J].�й���ҽ��ѧ, 2018 (1) :87-92.
����[22]���IJ�, ������, ������, ��.HIF-1α���߷���Ԥ�����������Ƥ����Ԫ�����е�����:��Slit2?Robo�ź�ͨ·�Ĺ�ϵ[J].�л�����ѧ��־, 2015, 35 (5) :550-554.
����[23]Ф����, �׳���, �°���, ��.��������������Slit2�ڳ�������Լ������˺�ı���[J].�л�ʵ�������־, 2006, 23 (8) :987-989.
����[24]������, ������, �ﲨ, ��.����Ƥ�����ν���ϸ�����ǰ����Slit2��Robo1�ı����о�[J].�з���������־, 2012, 29 (1) :4-8.
����[25]������, ����, ����.Slit2�ڲ�����ϵͳ�����е��о�[J].����������־, 2017, 25 (5) :317-319.
����[26]����Ƽ.��ͬ��������������Ĥ��ϸ����������GFAP��VEGF�Ķ�̬����[J].������������, 2017, 11 (7) :522-523.
����[27]�¾�, ��ΰ��, ���.��ϸ���ֻ�ʾ����GFAP����������ӫ�ⱨ��ϵͳ�ļ�ֵ[J].�й���֯�����о�, 2017, 21 (21) :3370-3375.
����[28]��, ���, Ф��, ��.��״�����Ѫ��NSEˮƽ������ʹ��������о�[J].�й���ʹҽѧ��־, 2017, 23 (7) :538-540.





