摘 要: 下丘脑背内侧核 (Dorsomedial Hypothalamic Nucleus, DMH) 是自主神经系统的重要组成部分, 同时与调控焦虑情绪的边缘系统存在广泛联系。然而, DMH调控焦虑等负性情绪的细胞机制目前尚不明确。该文应用光遗传技术特异性调控小鼠的谷氨酸能神经元和γ-氨基丁酸能神经元发现, 兴奋DMH的谷氨酸能和γ-氨基丁酸能神经元均能显着诱发焦虑样行为和心率上升, 抑制两种神经元均能抑制焦虑样行为并降低心率。该研究证实了DMH特定类型的神经细胞对焦虑的双向调控作用, 并有望为神经精神疾病的临床干预提供新的调控靶点。
关键词: 光遗传学; 下丘脑背内侧核; 焦虑; 心率;
Abstract: Dorsomedial hypothalamic nucleus (DMH) is an important component of autonomic nervous system, which has extensive connections with limbic system. DMH may participates in the regulation of negative emotions, such as anxiety and fear. However, the cellular mechanisms are still unknown. In this study, elevated plus maze and open filed tests were performed, and a bidirectional effect of optogenetic modulation on behaviors was found. Activating the DMH glutamatergic or gamma-aminobutyric acid (GABAergic) neurons leading to the anxiety-like behavior and increasing of heart rate, while optical inhibition significantly suppressed the anxiety-like behavior and decreased heart rate. These findings may provide new targets for clinical therapy for neuropsychiatric disorders.
Keyword: optogenetics; dorsomedial hypothalamic nucleus (DMH) ; anxiety; heart rate;
1、 引言
随着现代生活节奏不断加快, 人们工作和学习压力逐渐增大, 极易诱发情绪异常。焦虑是最常见的负性情绪之一, 对人们身心健康有着较大的危害。长时间焦虑易导致失眠[1], 诱发产生躁狂、抑郁、药物成瘾、精神分裂等精神疾病[2,3], 并增加罹患心脏病和癌症的风险[4,5]。因此, 解析产生负性情绪背后的神经回路机制, 揭示焦虑情绪的调控模式, 对于改善人情绪异常状况, 特别是长时间受焦虑情绪困扰的人群意义重大[6,7]。焦虑样行为通常被认为与腹侧海马 (Ventral Hippocampus) 、杏仁核 (Amygdala) 等脑区密切相关[8]。海马下托 (Subiculum) 作为海马的一个重要出口, 是介导大脑皮层 (Cerebral Cortex) 与皮层下脑区的重要核团。其中, 海马下托的γ-氨基丁酸 (GABA) 能系统和谷氨酸系统对于调控焦虑情绪起到重要的作用[9,10]。杏仁核由多个亚区组成, 其中的基底外侧杏仁核和中央杏仁核在编码焦虑样行为中的作用尤为显着[11,12]。首先, 外侧杏仁核接收来自丘脑 (Thalamus) 和前额叶皮层 (Prefrontal Cortex, PFC) 的感觉信息传入;其次, 信息流途经基底外侧杏仁核处理后, 再经过杏仁核基核并做进一步处理;最后, 信息流到达中央外侧杏仁核, 并在其中进行最终的处理编码[13,14]。
研究显示, 下丘脑也参与了对焦虑和恐惧等负性情绪的调节。下丘脑背内侧核 (Dorsomedial Hypothalamic Nucleus, DMH) 、下丘脑的腹侧内侧核团 (Ventromedial Hypothalamus, VMH) 、下丘脑前端乳头体 (Premamillary Region of the H y p o t h a l a m u s) 、下丘脑前端核团 (A n t e r i o r Hypothalamic Nucleus) 、下丘脑外侧核团 (Lateral Hypothalamus) 等的GABA能系统参与了恐惧的调控作用[15,16]。其中, 前额叶皮层和DMH的GABA能回路调节场景相关的本能学习与本能情绪[17]。前额叶皮层与内侧下丘脑存在广泛连接, 且通过NMDA和AMPA/Kainate谷氨酸受体与内侧下丘脑中GABA受体的相互作用调节防御行为和先天性焦虑诱导的回避伤害行为[18]。人们通过刺激DMH、VMH的背内侧区以及背侧导水管周围灰质 (Medial Pericatheter Gray) 发现, 这些核团与定向和非定向逃逸行为存在相关性[19]。此外, VMH和DMH的GABA受体阻滞引起焦虑样的行为, 并且产生先天性焦虑诱导的回避伤害行为[20,21]。
需要指出的是, DMH作为自主神经系统的较高级中枢, 与上述焦虑情绪相关的脑区存在广泛的投射关系。DMH与中央外侧杏仁核、中央内侧杏仁核、导管周围灰质、终纹床核 (Bed Nucleus of Stria Terminalis) 以及迷走神经背复合体 (Dorsal Vagal Complex) 等多种焦虑相关脑区存在投射关系, 并对其具有靶向调节作用[22]。越来越多研究证实, DMH在调节着体温、摄食、饮水、血糖和心率等重要生理功能的同时[23], 也参与对焦虑等负性情绪的调节。例如, 电刺激DMH和VMH会导致防御性行为的产生, 以及血压上升、皮肤及小肠血管收缩、心率加速等交感神经性反应[24]。DMH的Y神经肽影响内脏对胰岛素的敏感性, 进而会产生对焦虑的敏感样行为[23]。刺激DMH谷氨酸能中间神经元引起的心率失常[25]。DMH的GABA能神经元损伤引起的焦虑样行为[17]。DMH的5-羟色胺1A受体介导T迷宫升高时的恐慌相关反应[26]。然而, DMH调控焦虑等负性情绪的细胞机制目前尚不明确。
为此, 本研究采用光遗传技术来解析DMH中不同类型神经元对焦虑的影响。借助高架十字迷宫 (Elevated Plus-Maze Test, EPM) 和旷场 (Open Field Test, OFT) 行为学范式对小鼠的焦虑情绪的变化进行评估, 并同时监测小鼠的心率变化。我们在小鼠DMH中表达了不同类型的标记蛋白 (AAV9-CaMKIIα-ChR2-mcherry、AAV9-CaMKIIα-NpHR-mcherry、AAV9-DIO-ChR2-mcherry和AAV9-DIO-NpHR-mcherry) , 利用光纤植入体特异性激活或抑制DMH特定类型的神经元, 实现了对小鼠焦虑情绪的双向调节。本研究有望为临床干预焦虑症提供新的靶点和理论依据。
2、 实验
2.1、 实验前准备
2.1.1、 溶液的配制
磷酸盐缓冲液 (Phosphate Buffer Solution, PBS) 采用PBS缓冲液干粉 (品牌Solarbio) , pH为7.2~7.4。多聚甲醛溶液采用多聚甲醛粉末 (MOLBASE) , 按照4%的质量与体积比, 在60℃恒温条件下缓慢溶解于PBS缓冲液中配制而成。蔗糖溶液采用蔗糖粉末 (国药集团上海化学试剂有限公司) , 按照30%的质量与体积比, 在常温条件下溶解于PBS缓冲液中配制而成。水合氯醛麻醉溶液采用水合氯醛粉末 (国药集团化学试剂有限公司) , 按照10%的质量与体积比, 在常温条件下溶解于生理盐水 (国药集团化学试剂有限公司) 中配制而成, 并保存于棕色玻璃瓶中。上述溶液均保存于4℃冰箱中。
2.1.2、 光纤植入体的制备
光纤植入体采用市售的直径200μm光纤 (纽顿控股有限公司) 和与之匹配的陶瓷插芯 (纽顿控股有限公司) 制作而成。首先, 剪取适当长度的光纤, 用光纤剥线钳 (纽顿控股有限公司) 剥去一端的保护皮层 (1 cm左右) , 用钨钢刀手工环切端口处, 在光学显微镜下检查切口, 反复环切直到平整为止。然后, 光纤去皮端穿过陶瓷插芯, 在光纤的光出射端涂抹环氧树脂胶固定。用细砂纸轻轻打磨光纤的激光入口端, 显微镜下检查光纤与陶瓷插芯耦合整齐即可。最后, 根据脑区的深度环切光纤头的光出射端, 用市售商用的光强计 (Sanwa LP1手持式激光功率计) 测试光纤的透光率, 透光率达到60%以上的视为合格。
2.2、 实验动物
实验所用的小鼠均饲养在SPF级的专门饲养间内, 给予12 h-12 h交替与昼夜同步的光照条件。动物实验方案经中国科学院深圳先进技术研究院动物伦理委员会审查通过。其中, C57小鼠 (8~12周龄、雄性, 体重20 g左右) 购自北京维通利华实验动物技术有限公司;GAD-cre转基因小鼠 (8~12周龄、雄性, 体重20 g左右) 是中国科学院深圳先进技术研究院培育出的转基因品系, 经基因鉴定合格。
2.3、 手术实验
2.3.1、 病毒的注射
首先, 腹腔麻醉小鼠 (每10 g小鼠注射0.04 m L10%的水合氯醛麻醉溶液) , 待小鼠完全麻醉后, 用弯头剪刀 (上海金钟医疗器械有限公司) 剪去头毛, 露出头皮, 固定在立体定位仪 (瑞沃德) 上, 用碘伏消毒后, 剪开头皮并用手术镊 (FST, 5号) 取少许棉花擦拭头皮, 直到能够清晰看到颅骨骨缝为止。其次, 以前囟 (Bregma点) 和后囟 (Lambda点) 为参考点, 反复调节, 使得小鼠脑在前后、左右均处于水平位置。最后, 调节立体定位仪, 用微量注射器的针头确定注射目标脑区, 接着用颅骨钻打孔 (瑞沃德微型手持式颅钻) , 紧接着用注射泵 (瑞沃德微量注射泵Legato130) 吸取病毒 (吸取速率为100 nL/s) , 预注射50 nL, 确定注射针未堵塞 (看针头处有小液泡) 后, 缓慢地将针头旋转下降至目标脑区并开始注射 (注射速率100 nL/min) 。随后, 停针与注射相同的时间, 缓慢地将注射针头旋出脑组织, 并将小鼠伤口缝合消毒。
本实验对C57小鼠注射AAV9-CaMKIIα-ChR2-mcherry病毒, 对GAD-cre转基因小鼠注射AAV9-DIO-ChR2-mcherry病毒;作为对照组, 对C57小鼠注射AAV9-CaMKIIα-NpHR-mcherry病毒, 对GAD-cre转基因小鼠注射AAV9-DIO-NpHR-mcherry病毒。注射位点均为DMH (AP:-1.6, ML:-0.35, DV:-5.0) , 注射剂量均为300 nL。其中, 以小鼠脑前囟 (Bregma点) 为坐标原点、后囟 (Lambda点) 为 (0, 0, 4.24) 参考点建立空间直角坐标系。因此, AP代表小鼠脑的头尾段坐标;ML代表小鼠脑偏离中线的坐标;DV代表小鼠脑垂直坐标。
2.3.2、 埋植光纤植入体
首先, 重复2.3.1手术实验至擦拭清晰看到颅骨骨缝。在颅骨上选取适当的位置, 颅钻打孔后拧入颅钉 (4颗) 。其次, 以前囟 (Bregma点) 和后囟 (Lambda点) 为参考点, 反复调节, 使得小鼠脑在前后、左右均处于水平位置。最后, 调节立体定位仪, 并用光纤尖端确定注射目标脑区, 用颅钻打孔后, 再次调节立体定位仪, 缓慢地将光纤头旋转下降至目标脑区, 用牙科水泥封装光纤植入体, 待牙科水泥凝固后, 先松开定位仪上的光纤植入体, 再松开固定小鼠的耳杆。
本实验中, 所有实验小鼠光纤埋置时间为病毒注射后四周, 光纤埋置的位点均为DMH (AP:-1.6, ML:-0.35, DV:-4.8) 。为了使结果具有较好的光刺激效果, 光纤的埋植位点要比病毒注射位点浅0.2 mm。
2.3.3、 灌流及脑组织的固定过程
首先, 将完全麻醉后的小鼠腹部朝上固定在灌流台上, 用手术剪刀剪开小鼠的腹腔和胸腔, 在左心室心尖处插入灌流针头, 用止血钳固定, 剪破右心房, 先后灌注PBS溶液 (20 mL/min) 和PFA固定溶液 (18 mL/min) 。其次, 完成灌流后, 将脑组织取出并浸泡在4%的PFA固定液中固定, 随后更换为30%的蔗糖溶液。最后, 当观察到小鼠脑沉到蔗糖溶液的底部时, 取出包埋并存放在-20℃冰箱中。
2.4、 切片与免疫组化分析
首先, 将包埋后的鼠脑固定, 用冰冻切片机切片 (Leica CM1950冰冻切片机) 。其中, 切片厚度为30μm。然后, 收集目标脑区脑片, 将其浸泡于NaN3-PBS溶液 (NaN3, 0.1%) , 洗涤 (PBS溶液) 并贴脑片, 滴加4′, 6-二脒基-2-苯基吲哚 (DAPI) 荧光染料, 加盖盖玻片后封片。最后, 玻片扫描 (Olympus全切片扫描仪) 拍照, 观察目标脑区荧光标记情况, 进而确定病毒注射和光纤埋植位点。
2.5、 行为学和心率测试实验
2.5.1、 行为学测试装置
本研究采用光遗传调控技术, 结合高架十字迷宫 (EPM) 和旷场 (OFT) 来研究DMH与焦虑情绪之间的关联。
实验所用EPM和OFT装置材质均为白色无味的塑料材质, 均参照国际上的通用规格设计 (EPM:高度为60 cm, 臂长为56 cm, 臂宽为6 cm;OFT:底边边长为50 cm, 四壁高度为60 cm) 。
2.5.2、 光遗传刺激参数
EPM、OFT和心率测试实验的光遗传刺激参数均相同。波长, 473 nm蓝光 (激活实验) 、589 nm黄光 (抑制实验) ;刺激频率, 20 Hz (C57小鼠) 、40 Hz (GAD-cre转基因小鼠) ;延迟, 0.0 ns;幅度, 5.0峰峰值电压;直流分量, 0 mv;斜率, 正;脉冲宽度, 5.0 ms;触发延迟, 0.0 ns。调节激光器的输出, 当光纤末端功率为10 mW/mm2左右时为合格刺激光源。
2.5.3、 高架十字迷宫 (EPM) 测试实验
开始测试时, 将实验动物放入高架十字迷宫中央区, 头朝开放臂方向。每只实验动物测试时的放置位点和头部朝向均保持一致。打开摄像机, 以5 min无光刺激、5 min光刺激、5 min无光刺激的模式拍摄, 记录动物在开放臂、闭合臂和中央区的行为。实验全过程中, 实验人员距离高架十字迷宫保持1 m的距离。每只实验动物行为记录结束之后放回原鼠笼内, 同时清理十字迷宫, 接着用5%的醋酸水溶液或20%的酒精擦拭迷宫内部, 消除上一只动物残留气味对后继实验动物的影响。
2.5.4、 旷场 (OFT) 测试实验
开始测试时, 将实验动物放入旷场中央区, 每只实验动物测试时的放置位点和头部朝向均保持一致。打开摄像机, 以10 min无光刺激、10 min光刺激、10 min无光刺激的模式拍摄, 记录动物在中央区和边缘区的行为。实验全过程中, 实验人员距离旷场装置保持1 m的距离。每只实验动物拍摄记录结束之后放回原鼠笼内, 同时清理旷场装置, 接着用5%的醋酸水溶液或20%的酒精擦拭旷场内部, 消除上一只实验动物残留的气味对后继实验动物的影响。
2.5.5、 心率测试实验
动物在遭遇焦虑、恐惧等负性情绪来袭时, 生理信号, 如呼吸频率、心率、血压以及血氧饱和度等生理信号会发生相应的变化。我们在浅麻醉 (每10 g小鼠注射0.02 mL 10%的水合氯醛麻醉溶液) 状态下测试动物在接受光刺激前后的心率变化, 作为其焦虑情绪变化的一个补充证据。实验采用脉搏血氧仪 (Starr life science, MouseOx) , 在无创条件对生理信号进行检测和记录。待小鼠微麻醉后, 用脱毛膏对小鼠颈部进行脱毛处理, 之后将小鼠放在单独的笼具内, 夹上脖圈 (生理信号的输入装置) , 待信号稳定后, 开始记录测量。整个测量过程中, 保持小鼠的颈部伸展, 防止颈部弯曲窒息。每只老鼠测试5~8个循环, 每个循环包含如下阶段:60 s无光刺激、30 s光刺激、60 s无光刺激。每个循环都要待信号稳定后再开始测量, 且全过程保持信号稳定。
2.6、 数据分析
2.6.1、 行为学数据分析
行为学视频均利用ANY-maze动物行为分析系统 (美国Stoelting公司开发) 自动分析统计。本研究EPM统计光刺激前后, 即5 min+5 min+5 min, 动物进入开放臂的次数、进入开放臂的时间;OFT统计光刺激前后, 即10 min+10 min+10 min, 动物进入中央区次数、进入中央区的时间。软件根据预先划定的区域将自动把这些参数分类统计得到动物活动情况报告表格, 并绘制小鼠的行为学数据图。
2.6.2、 心率数据分析
统计每只小鼠在60 s+30 s (无光刺激+光刺激) 一个循环中各个阶段心率的平均值, 然后选取5个循环, 再统计同一只小鼠在不同循环中对应阶段心率均值的均值, 导出数据统计表格, 并绘制小鼠的心率变化图。
3、 实验结果
3.1 、光遗传兴奋C57小鼠DMH谷氨酸能神经元
为了研究DMH谷氨酸能神经元对焦虑情绪是否存在调控作用, 我们将AAV9-CaMKIIα-Ch R2-mcherry病毒蛋白选择性地表达在了C57小鼠的DMH区域 (图1 (a) ) 。图1 (b) 显示, 光遗传兴奋DMH区域的谷氨酸能神经元能导致小鼠心率的上升 (未刺激时心率: (482.85±23.37) bpm, 刺激时心率: (5 2 3.3 4±2 6.6 9) b p m, mean±SD (均值±标准差) , P=0.042 08, 每组n=8) , 表明光刺激影响了小鼠的生理状态。此外, 在EPM实验中 (图1 (c-d) ) , 光刺激时小鼠进入开放臂的次数减少 (未刺激时进入开放臂的次数:6.57±2.89, 刺激时进入开放臂的次数:1.28±1.46, mean±SD, P=0.000 91, 每组n=5) 且进入开放臂的时间减少 (未刺激时进入开放臂的时间: (38.98±13.86) s, 刺激时进入开放臂的时间: (4.58±2.54) s, mean±SD, P=0.000 87, 每组n=7) 。在OFT测试实验中 (图1 (e-f) ) , 光刺激时小鼠进入中央区域的次数减少 (未刺激时进入中央区的次数:8.45±3.56, 刺激时进入中央区的次数:3.35±1.25, mean±SD, P=0.002 98, 每组n=6) 且进入中央区域的时间减少 (未刺激时进入中央区的时间: (30.02±5.63) s, 刺激时进入中央区的时间: (10.23±2.32) s, mean±SD, P=0.005 96, 每组n=7) 。上述结果表明, 光遗传兴奋C57小鼠DMH的谷氨酸能神经元后, 小鼠心率上升且表现出焦虑样行为。
3.2、 光遗传抑制C57小鼠DMH谷氨酸能神经元
作为对照实验, 我们将AAV9-CaMKIIα-Np HR-mcherry病毒蛋白选择性地表达在了C57小鼠的DMH区域 (图2 (a) ) 。图2 (b) 显示, 光遗传抑制DMH区域的谷氨酸能神经元能导致小鼠心率的下降 (未刺激时心率: (465.82±13.21) bpm, 刺激时心率: (413.34±21.62) bpm, mean±SD, P=0.049 88, 每组n=8) , 表明光刺激影响了小鼠的生理状态。此外, 在EPM实验中 (图2 (cd) ) , 光刺激时小鼠进入开放臂的次数增加 (未刺激时进入开放臂的次数:2.872±2.68, 刺激时进入开放臂的次数:4.02±2.36, mean±SD, P=0.040 91, 每组n=6) 且进入开放臂的时间增加 (未刺激时进入开放臂的时间: (8.98±1.82) s, 刺激时进入开放臂的时间: (20.01±5.54) s, mean±SD, P=0.008 87, 每组n=6) 。在OFT测试实验中 (图2 (e-f) ) , 光刺激时小鼠进入中央区域的次数增加 (未刺激时进入中央区的次数:5.15±3.53, 刺激时进入中央区的次数:9.36±3.28, mean±SD, P=0.009 08, 每组n=6) 且进入中央区域的时间增加 (未刺激时进入中央区的时间: (8.42±5.69) s, 刺激时进入中央区的时间: (20.56±4.31) s, mean±SD, P=0.008 96, 每组n=6) 。上述结果表明, 光遗传抑制C57小鼠DMH的谷氨酸能神经元后, 小鼠心率下降且表现出焦虑下降行为。
图1 C57小鼠在光遗传兴奋DMH谷氨酸能神经元前后的心率和行为变化 (*P0.05, **P0.01, ***P0.001, 配对t-检验)
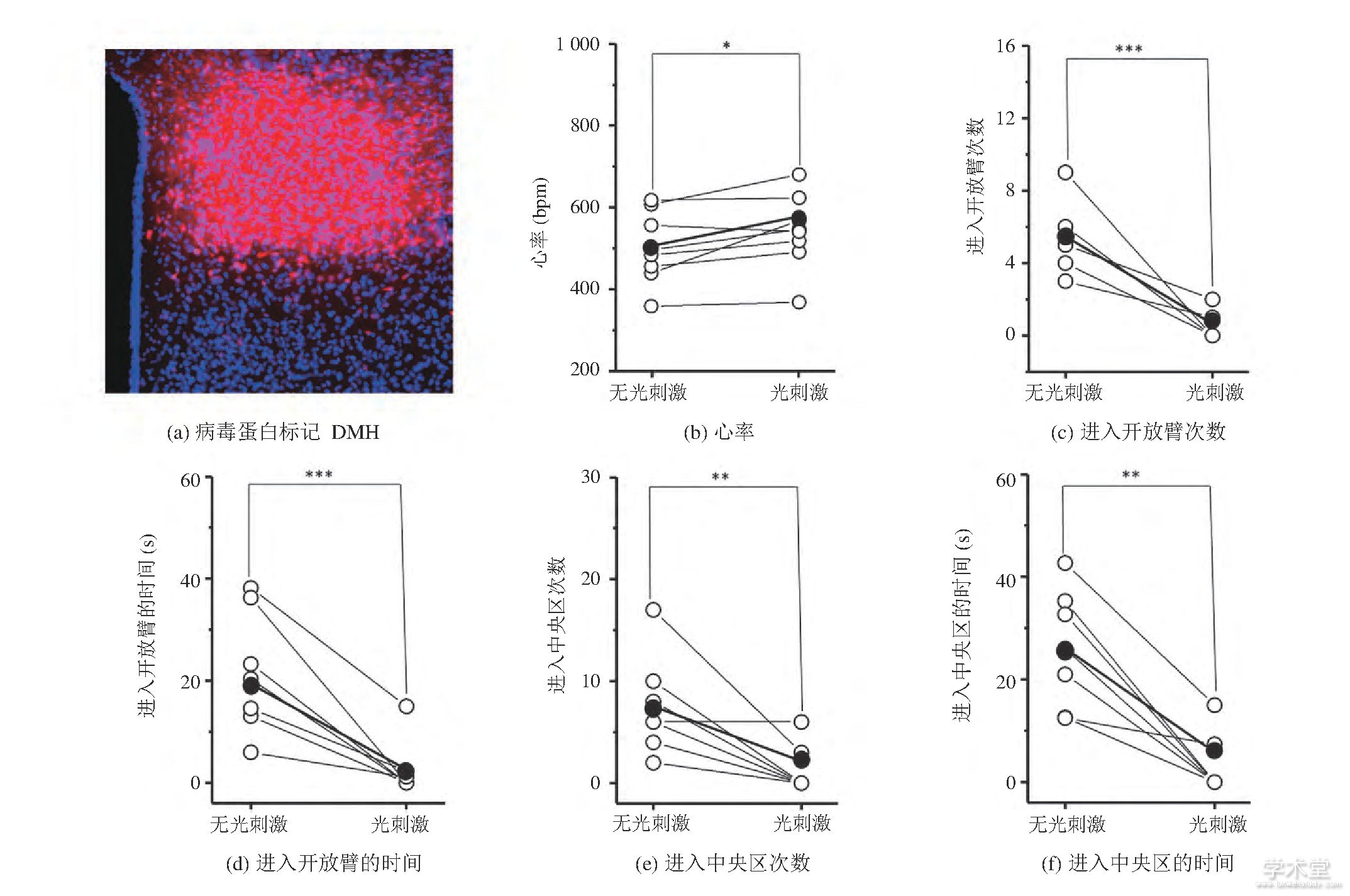
Fig.1 Heart rate and behavior changes of C57 mice before and during optogenetic excitation of DMH glutamatergic neurons (*P<0.05, **P<0.01, ***P<0.001, Paired t-Test)
注:图 (b-f) 中, 每个空心球代表每只小鼠的参数, 实心球代表参数均值
3.3、 光遗传兴奋GAD-cre小鼠DMH的GABA能神经元
为了研究DMH GABA能神经元对焦虑情绪是否存在调控作用, 我们将AAV9-DIO-ChR2-mcherry病毒蛋白选择性地表达在了GAD-cre小鼠的DMH区域 (图3 (a) ) 。图3 (b) 显示, 光遗传兴奋DMH区域的GABA能神经元能导致小鼠心率的上升 (未刺激时心率: (510.86±25.36) bpm, 刺激时心率: (573.72±24.64) bpm, mean±SD, P=0.047 58, 每组n=8) , 表明光刺激影响了小鼠的生理状态。此外, 在EPM实验中 (图3 (cd) ) , 光刺激时小鼠进入开放臂的次数减少 (未刺激时进入开放臂的次数:13.55±2.86, 刺激时进入开放臂的次数:6.27±1.96, mean±SD, P=0.007 91, 每组n=7) 且进入开放臂的时间减少 (未刺激时进入开放臂的时间: (36.98±12.86) s, 刺激时进入开放臂的时间: (19.58±6.54) s, mean±SD, P=0.008 87, 每组n=7) 。在OFT测试实验中 (图3 (e-f) ) , 光刺激时小鼠进入中央区域的次数减少 (未刺激时进入中央区的次数:8.95±3.56, 刺激时进入中央区的次数:5.34±3.25, mean±SD, P=0.044 98, 每组n=6) 且进入中央区域的时间减少 (未刺激时进入中央区的时间: (25.13±6.63) s, 刺激时进入中央区的时间: (9.23±3.31) s, mean±SD, P=0.009 96, 每组n=7) 。上述结果表明, 光遗传兴奋GAD-cre小鼠DMH的GABA能神经元后, 小鼠心率上升且表现出焦虑样行为。
图2 C57小鼠在光遗传抑制谷氨酸能神经元前后的心率和行为变化 (*P0.05, **P0.01, 配对t-检验)
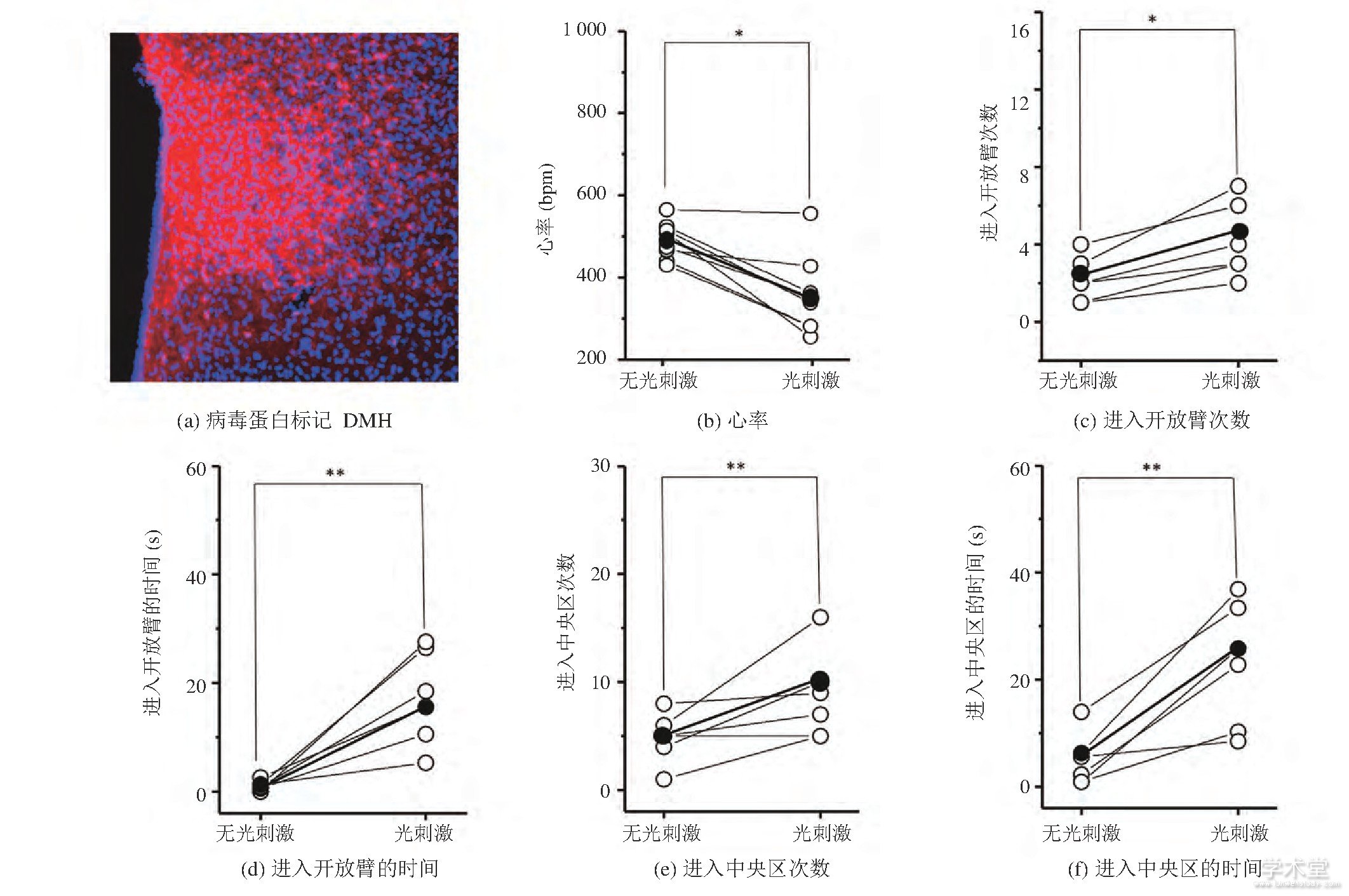
Fig.2 Heart rate and behavior changes of C57 mice before and during optogenetic inhibition of DMH glutamatergic neurons (*P<0.05, **P<0.01, Paired t-Test)
注:图 (b-f) 中, 每个空心球代表每只小鼠的参数, 实心球代表参数均值
3.4、光遗传抑制GAD-cre小鼠DMH的GABA能神经元
作为对照实验, 我们将AAV9-DIO-NpHR-mcherry病毒蛋白选择性地表达在了GAD-cre小鼠的DMH区域 (图4 (a) ) 。图4 (b) 显示, 光遗传抑制DMH区域的GABA能神经元能导致小鼠心率的下降 (未刺激时心率: (480.86±9.98) bpm, 刺激时心率: (423.39±8.69) bpm, mean±SD, P=0.046 78, 每组n=6) , 表明光刺激影响了小鼠的生理状态。此外, 在EPM实验中 (图4 (cd) ) , 光刺激时小鼠进入开放臂的次数增加 (未刺激时进入开放臂的次数:2.48±2.62, 刺激时进入开放臂的次数:3.06±2.86, mean±SD, P=0.049 91, 每组n=6) 且进入开放臂的时间增加 (未刺激时进入开放臂的时间: (5.78±1.52) s, 刺激时进入开放臂的时间: (16.71±6.58) s, mean±SD, P=0.009 85, 每组n=6) 。在OFT测试实验中 (图4 (e-f) ) , 光刺激时小鼠进入中央区域的次数增加 (未刺激时进入中央区的次数:4.27±2.93, 刺激时进入中央区的次数:8.65±3.78, mean±SD, P=0.008 48, 每组n=6) 且进入中央区域的时间增加 (未刺激时进入中央区的时间: (10.77±4.43) s, 刺激时进入中央区的时间: (23.96±8.71) s, mean±SD, P=0.008 56, 每组n=6) 。上述结果表明, 光遗传抑制GAD-cre小鼠DMH的GABA能神经元后, 小鼠心率下降且表现出焦虑下降行为。
图3 GAD-cre小鼠在光遗传兴奋DMH GABA能神经元前后的心率和行为变化 (*P0.05, **P0.01, 配对t-检验) Fig.3 Heart rate and behavior changes of GAD-cre mice before and during optogenetic excitation of DMH GABAergic neurons (*P<0.05, **P<0.01, Paired t-Test)
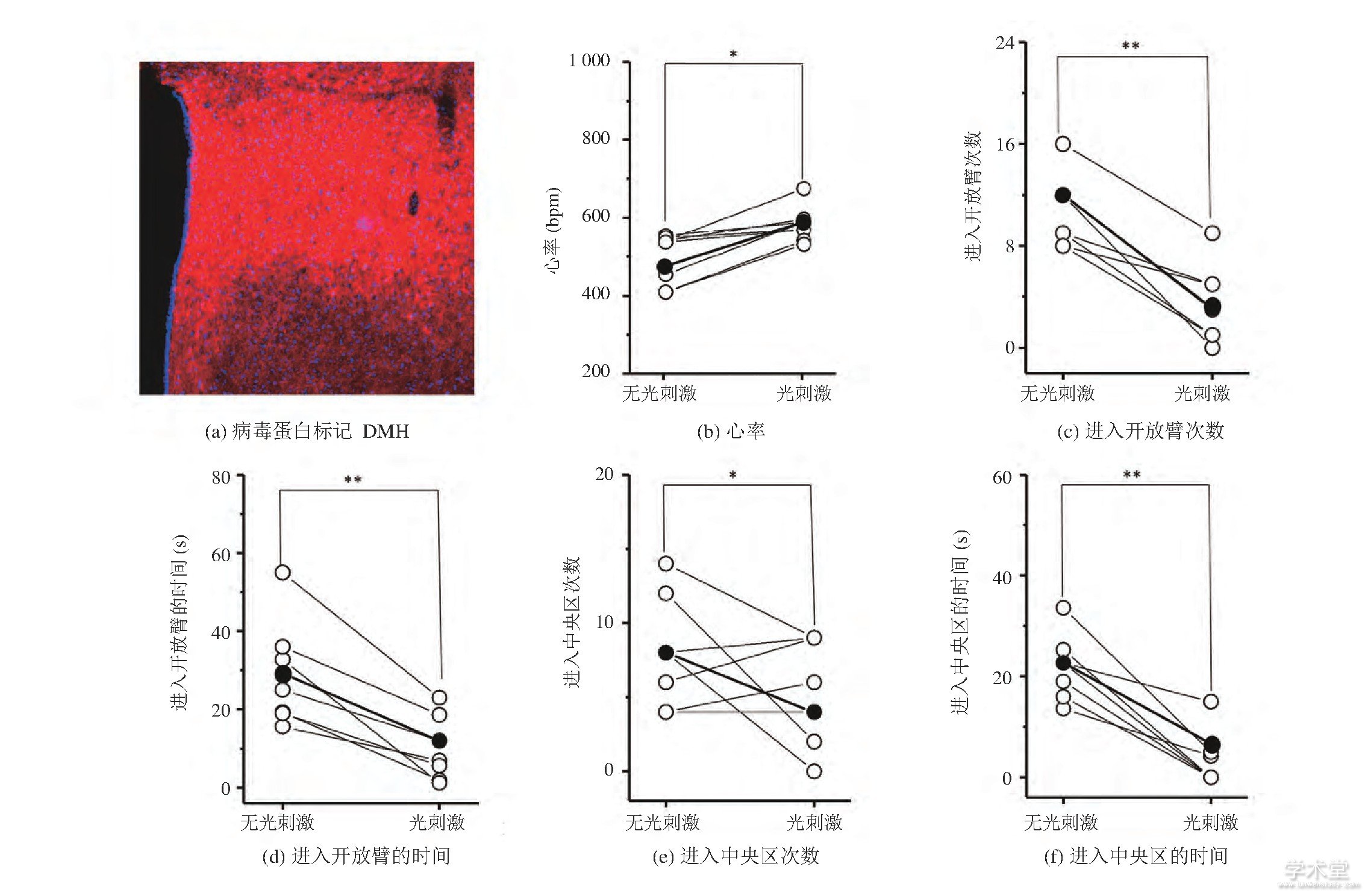
注:图 (b-f) 中, 每个空心球代表每只小鼠的参数, 实心球代表参数均值
4、 与国内外相似研究对比分析
关于DMH, 通常人们将其作为自主神经系统的重要部分来研究, 认为DMH直接或间接控制内脏和内分泌腺体, 进而调节性行为、打斗、心率、血压、体温等至关重要的生理功能, 任何微小下丘脑的病变或损伤都会引起诸多生理活动与功能的紊乱和异常[23]。López-González等[27]用电刺激麻醉大鼠下丘脑防御区诱发心血管加速, 而在此过程脑桥A5区神经元全程参与其中。这一研究结论说明脑桥A5区的神经元可能参与了由下丘脑防御区引起的心血管反应机制。SévozCouche等[28]发现下丘脑DMH参与了慢性心血管疾病的调控。但仍较少研究将下丘脑的活动与焦虑、紧张、愤怒、沮丧、悲伤、痛苦等负性情绪的变化联系在一起[15]。本文的一大创新点就是将DMH与调控焦虑情绪的经典脑区联系在一起研究, 结果发现DMH参与了焦虑的调控机制, 并对DMH进行了细胞分类。
图4 GAD-cre小鼠在光遗传抑制DMH GABA能神经元前后的心率和行为变化 (*P0.05, **P0.01, 配对t-检验)
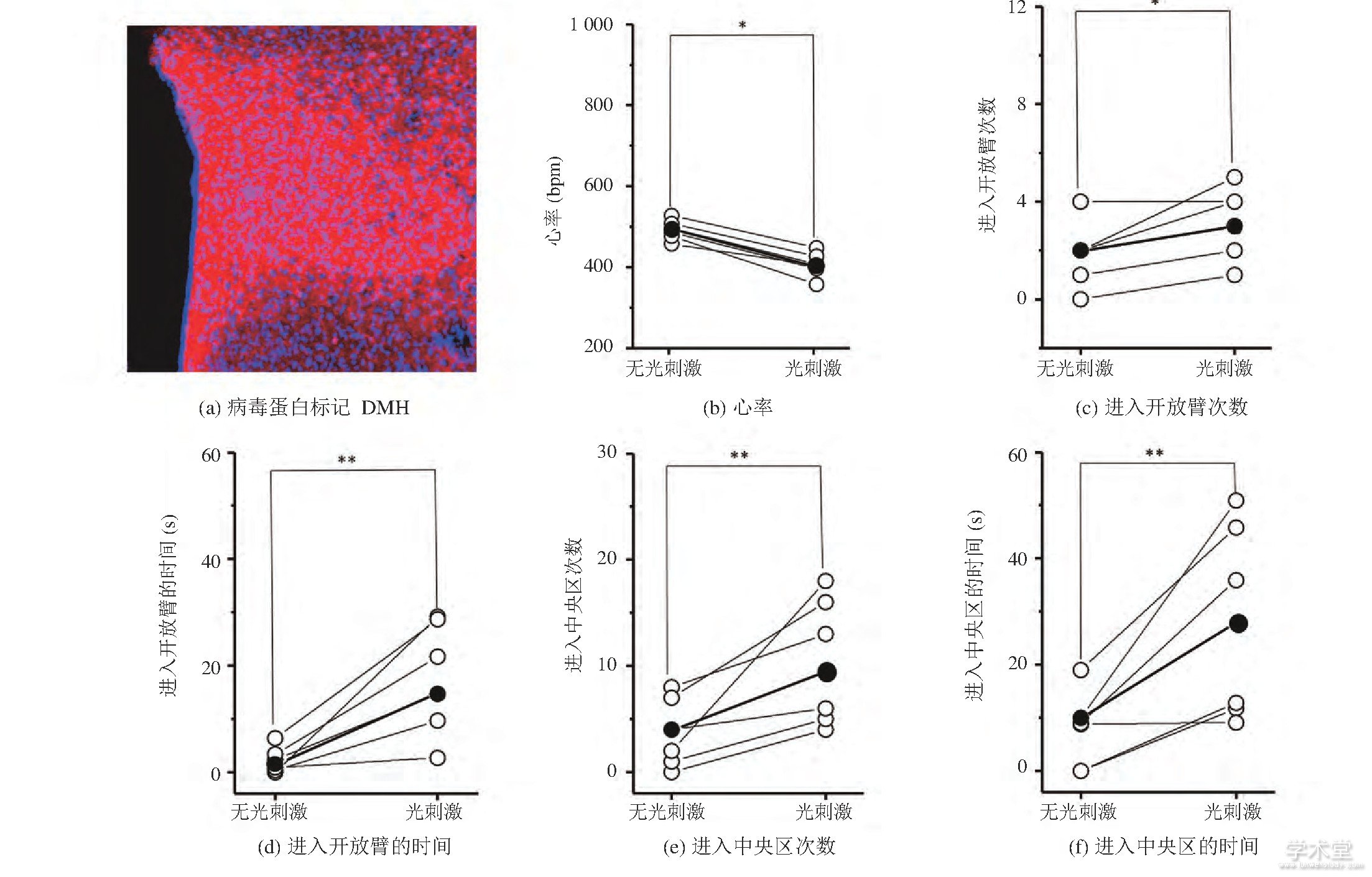
Fig.4 Heart rate and behavior changes of GAD-cre mice before and during optogenetic inhibition of DMH GABAergic neurons (*P<0.05, **P<0.01, Paired t-Test)
注:图 (b-f) 中, 每个空心球代表每只小鼠的参数, 实心球代表参数均值
另外, 与传统的电刺激DMH的研究方法相比, 本文采用的光遗传技术能在活体动物甚至是自由运动的动物脑内, 精准地调控特定种类神经元的活动。其中, 在时间上的精确度可达到毫秒级别, 在空间上的精确度则能达到单个细胞级别, 并且调控具有细胞选择性[29,30]。
5、 总结
为了研究DMH中不同类型的神经元在负性情绪, 尤其是焦虑中的作用, 本文结合光遗传技术、生理参数检测技术和行为学方法, 对DMH内GABA能神经元、谷氨酸能神经元的正向激活与反向抑制进行特异性调控, 实现了对焦虑情绪的双向调控。
研究发现, 光遗传兴奋C57小鼠DMH的谷氨酸能神经元, 小鼠表现出对高架十字迷宫开放臂和旷场中央区域更少的探索行为。与此同时, 小鼠心率伴随光刺激而有所上升, 光停止刺激后, 心率恢复到刺激前的水平。结果提示小鼠行为上表现出焦虑上升, 与此同时, 生理上伴随着心率升高。反之, 若光遗传抑制C57小鼠DMH的谷氨酸能神经元, 小鼠表现出对高架十字迷宫开放臂和旷场中央区域更多的探索行为。同时小鼠心率伴随光刺激而有所下降, 光停止刺激后, 心率恢复到刺激前的水平。说明小鼠行为上表现出焦虑下降, 在生理上伴随着心率下降。这一结论表明, DMH的谷氨酸能神经元可能参与了小鼠焦虑样情绪的调控, 而且参与了自主神经系统对心率等生理信号的调控。
出乎意料的是, 光遗传兴奋GAD-cre小鼠DMH的GABA能神经元, 小鼠表现出显着的焦虑样行为和心率升高。反之, 若光遗传抑制GAD-cre小鼠DMH的GABA神经元, 小鼠在行为上表现出焦虑下降, 生理上伴随着心率下降。这一发现提示, 与DMH的谷氨酸能神经元的作用相同, DMH的GABA能神经元也参与了小鼠焦虑样情绪和心率等生理信号的双向调控。
综上所述, 本研究发现激活DMH区域的谷氨酸能神经元和GABA能神经元均导致了显着的焦虑样行为和心率的显着上升。反之, 抑制这一区域的谷氨酸能神经元和GABA能神经元均引起了焦虑样行为和心率的显着下降。这一发现为DMH参与对焦虑的调控提供了直接的证据。然而, 对于DMH调控焦虑的确切神经环路机制, 以及自主神经调节与情绪调节之间的相互作用等问题, 仍有待进一步研究。由于下丘脑各个核团的投射广泛, 神经环路功能复杂, 本文针对DMH中谷氨酸能细胞、GABA能细胞两类细胞对心率和焦虑情绪的影响进行了初步的研究, 发现了两者的协同作用。本研究有望为将来解析诸多神经-精神共患病的调控机制提供可能的靶点和理论支持。
参考文献
[1]Tye KM, Deisseroth K.Optogenetic investigation of neural circuits underlying brain disease in animal models[J].Nature Review Neuroscience, 20123171 (13) :251-266.
[2]Lu Y, Zhong C, Wang LL, et al.Optogenetic dissection of ictal propagation in the hippocampalentorhinal cortex structures[J].Nature Communications, 2016, 10962 (7) :1-11.
[3]Boido D, Jesuthasan N, de Curtis M, et al.Network dynamics during the progression of seizure-like events in the hippocampal-parahippocampal regions[J].Cereb Cortex, 2014, 24 (1) :163-173.
[4]Paz JT, Davidson TJ, Frechette ES, et al.Closedloop optogenetic control of thalamus as a tool for interrupting seizures after cortical injury[J].Nature Neuroscience, 2013, 16 (1) :64-70.
[5]Einevoll GT, Kayser C, Logothetis NK, et al Modelling and analysis of local field potentials for studying the function of cortical circuits[J].Nature Review Neuroscience, 2013, 3599 (14) :770-785.
[6]Yi zhar O, FennoL F, Davidson T J, et al Optogenetics in neural systems[J].Neuron, 2011, 71 (1) :9-34.
[7]Goldberg EM, Coulter DA.Mechanisms of epileptogenesis a convergence on neural circuit dysfunction[J].Nature Review Neuroscience2013, 3482 (14) :337-349.
[8]Wen JL, Xue L, Wang RH, et al.Involvement of the dopaminergic system in the consolidation of fear conditioning in hippocampal CA3 subregion[J].Behavioural Brain Research, 2015, 278:527-534.
[9]Tang H, Wu GS, Xie J, et al.Brain-wide map of projections from mice ventral subiculum[J]Neuroscience Letters, 2016, 629:171-179.
[10]Wang Y, Xu CL, Xu ZB, et al.Depolarized GABAergic signaling in subicular microcircuits mediates generalize dsei zureintem poral lobeepilepsy[J].Neuron, 2017, 95 (1) :92-105.
[11]Janak PH, Tye KM.From circuits to behaviour in the amygdala[J].Nature, 2015, 517 (7534) :284-292.
[12]Méndez-Bértolo C, Moratti S, Toledano R, et al.A fast pathway for fear in human amygdala[J]Nature Neuroscience, 2016, 19 (8) :1041-1049.
[13]Likhtik E, Stujenske JM, Topiwala MA, et al Prefrontal entrainment of amygdala activity signals safety in learned fear and innate anxiety[J].Nature Neuroscience, 2013, 17 (1) :106-113.
[14]McCullough KM, Choi D, Guo JD, et al.Molecular characterization of Thy1 expressing fear-inhibiting neurons within the basolateral amygdala[J].Nature Communications, 2016, 7:1-13.
[15]Gross CT, Canteras NS.The many paths to fear[J]Nature Review Neuroscience, 2012, 13 (9) :651-658.
[16]Santos JM, Macedo CE, Brand?o ML.Gabaergic mechanisms of hypothalamic nuclei in the expression of conditioned fear[J].Neurobiology of Learning and Memory, 2008, 90 (3) :560-568.
[17]Lissek S, Golisch A, Glaubitz B, et al.The GABAergic system in prefrontal cortex and hippocampus modulates context-related extinction learning[J].Brain Imaging and Behavior, 2017, 11 (16) :1885-1900.
[18]Silva Jr LG, de Menezes RCA, dos Santos RAS, et al.Role of periaqueductal gray on the cardiovascular response evoked by disinhibition of the dorsomedial hypothalamus[J].Brain Research2003, 984 (1-2) :206-214.
[19]Wong LC, Wang L, D’Amour JA, et al.Effective modulation of male aggression through lateral septum to medial hypothalamus projection[J].Current Biology, 2016, 26 (5) :593-604.
[20]Abreu AR, Molosh AI, Johnson PL, et al.Role of medial hypothalamic orexin system in panic, phobia and hypertension[J].Brain Research, 2018, doi:10.1016/j.brainres.2018.09.010.
[21]Zoeram SB, Salmani ME, Lashkarbolouki T, et al.Hippocampal orexin receptor blocking prevented the stress induced social learning and memory deficits[J].Neurobiology of Learning and Memory2019, 157:12-23.
[22]Root CM, Denny CA, Hen R, et al.The participation of cortical amygdala in innate, odour-driven behaviour[J].Nature, 2014, 515 (7526) :269-273.
[23]Li L, de La Serre CB, Zhang N, et al.Knockdown of neuropeptide Y in the dorsomedial hypothalamus promotes hepatic insulin sensitivity in male rats[J]Endocrinology, 2016, 157 (12) :4842-4852.
[24]Crick FH.Thinking about the brain[J].Scientific American, 1979, 241 (3) :219-232.
[25]Zhao S, Tang M, Yuan K, et al.Median nerve stimulation reduces ventricular arrhythmias induced by dorsomedial hypothalamic stimulation[J]Journal of Interventional Cardiac Electrophysiology2016, 47 (3) :275-283.
[26]Nascimento JOG, Kikuchi LS, Bortoli VC, et al.Dorsomedial hypothalamus serotonin 1A receptors mediate a panic-related response in the elevated T-maze[J].Brain Research Bulletin, 2014, 109:39-45.
[27]López-González MV, Díaz-Casares A, PeinadoAragonés CA, et al.Neurons of the A5 region are required for the tachycardia evoked by electrical stimulation of the hypothalamic defence area in anaesthetized rats[J].Experimental Physiology2013, 98 (8) :1279-1294.
[28]Sévoz-Couche C, Brouillard C, Camus F, et al Involvement of the dorsomedial hypothalamus and the nucleus tractus solitarii in chronic cardiovascular changes associated with anxiety in rats[J].The Journal of Physiology Neuroscience, 2013, 591 (7) :1871-1887.
[29]Silva BA, Mattucci C, Krzywkowski P, et al Independent hypothalamic circuits for social and predator fear[J].Nature Neuroscience, 201316 (12) :1731-1733.
[30]Silva BA, Mattucci C, Krzywkowski P, et al.The ventromedial hypothalamus mediates predator fear memory[J].European Journal of Neuroscience2016, 43 (11) :1431-1439.





