������Ѫ�ܼ���ȱ����������Ӧ������ȱ���Է�Ѫ�ܽṹ�ؽ���ȱ���Էζ�����ѹ��������Ҫ���ƣ���Ѫ���ع�����Ҫ�IJ����仯�Ƿζ���ƽ����ϸ��(pulmonary artery smooth muscle cells��PASMCs)����Ĥ����Ǩ������Ĥ�ϲ�������������ʧ�⡣�о�������ȱ���Էζ�����ѹѪ�ܽṹ�ؽ���Զ�˷ζ������ֵıȽ��˷ζ����������ţ������ڵ��������������Ѫ������ʵ���У�С������������������ԶԶ���ڴ������ɼ�С�����IJ����仯�ڼ�������������������Ҫ�����á�������ͬ���IJ����̼��£�С�������������ع��Ļ����в���ȷ����ˣ�̽��������֤��С������������ܵIJ��죬��������С�����������ع����ܵIJ������������ƣ�����Ԥ�����ڷ�Ѫ�ܵ��ع��Լ��ӻ��ζ�����ѹ�ķ�������չ������Ҫ�����о�����ԭ�������Ĵ���ͬ���� PASMCs��̽���˵����Բ�ͬ���� PASMCs ������Ǩ�Ƶ�Ӱ�졣
����1 �����뷽��
����1�� 1 ��Ҫ�Լ�������ƽ����ϸ������������ (dulbecco’s modifiedeagle’s medium��DMEM)�������� Sciencell ��˾;̥ţѪ��(fetal calf serum��FBS)�������� Gibcobrl ��˾;��ù�ء���ù�ؾ��������� Invitrogen ��˾;�ȵ���ø����������(dimethyl sulfoxide��DMSO)���Լ���������Sigma ��˾; EDTA���ļ�ż������ ( methyl thiazolyltetrazolium��MTT)�������� Amresco ��˾;PCNA �������� Santa Cruz Biotecnology;ƽ������������(α-smoothmuscle actin��α-SMA)����Ӣ�� Abcam ��˾;β-actin �������� Sigma ��˾�������Լ���Ϊ������������
����1�� 2 ʵ�鷽��
����1) ���� PASMCs ��ԭ������:ȡ������ԼΪ250 �� 300 g �������� SD ���� 4 ֻ������ҽ�ƴ�ѧ����ʵ�������ṩ��ʵ�鶯������֤��:SYXK(��)2010-0020����ǻע�� 2% ��ͱ�����(50 mg / kg) ��������������Ѹ�ٷ�����ķ���֯������ʢ�к��� 1%����ù�ص� PBS �������У���ʽ�����·������Ҷ�ķζ��������շζ����ķּ�����Ѫ�ܷ�Ϊ 3��:������(n =4)�������ζ����ɼ�����һ������;�ж�����(n =4)�������������ɵĶ�����������֧;С������(n =4)���������ڶ����ļ����Ϸ�֧�����ۿ���С�İ�����Ĥ����������ζ������ùγ�������� 2 �� 3 �飬ȥ����Ĥ��������Ĥƽ������֯�㣬���ۿƼ��������� 1 mm ×1 mm ��С��֯�飬������Ԥ��ʢ��3 mL ���ͽ�ԭø����Һ(1 800 U/mL)�����Ĺ��� 37 ��ˮԡ���� 20 min������֯��Ե������״��1 000 r / min ���� 5 min��������Һ������֯�����м��뺬 10%̥ţѪ��� DMEM �����������ᴵ���Ƴ�ϸ����Һ������ 35 mm �������У��� 5% CO2������ 37�� ������
����2) ʵ�����:ÿ��ʵ��ǰȡ״̬���õ� 4 �� 7 ������������ϸ������ϸ������������ 80% ��90% �ں�ʱ����ϸ�������������֡���ϸ�����ں�Ϊ��Ѫ�� DMEM ����������24h ʹϸ��ͬ�������ٲ������ֱ��������Ϊ:����������(DMEM + 2% FBS��5%CO2��21%O2��37 ������ 48 h);����������(DMEM +2% FBS��5% CO2��3%O2��37 ������ 48 h)��
����3) �� �� PASMCs �� �� �� ӫ �� �� ��: �� �� ��PASMCs ������ 24 �װ��ڵĸDz�Ƭ�ϣ������������Ѽ�ϸ����Ƭ��4%�Ķ�ۼ�ȩ���¹̶� 30 min������10% ��Ѫ���� 30 min��0�� 2% Triton X-100 ��Һ���·��� 30 min������һ��(anti-α-SMA��1�� 100)4 �������ҹ��Alexa Fluor 488 ��ǵĶ��� IgG (1�� 100)���±ܹ���� 1 h���ú� DAPI �ķ�Ƭ����Ƭ��۽�ӫ�������¹۲�����
����4)MTT �����:��ÿ�� 5 × 103��ϸ�������� 96�װ壬ÿ���� 6 �����ף��費��ϸ��ֻ������Һ�Ŀ�Ϊ�հס���ϸ�����ں������Ѫ�� DMEM ���������� 24 h ʹϸ��ͬ�������� 2 �飬�ֱ��ڳ����͵������������� 48 h����Ԥ������ÿ���� 200μL �����Ƶ� MTT(0�� 5 g/L)������ 37 �桢5% CO2�������ڼ������� 4 h����ȥ����Һ��ÿ���� 150 μL ��������(DMSO)���� 96 �װ�����ȫ����ø�����е����� 10 min��ѡ�� 490 nm ��������ø�����ϲⶨ���������ֵ��ʵ���ظ� 3 �Ρ�
����5)Western blotting ���: ȡ���鴦����Ĵ���PASMCs������������PBS ��ϴ 2 �飬����Ԥ��� ��IPA�����ѽ�Һ(��1%����ø���Ƽ�)30 μL �ѽ�ϸ����15min ���ռ��� 1�� 5 mL EP ���У��������� 3 �Σ����Ͼ��� 30 min �� 4 �桢12 000 r/min ���� 5 min����ȡ����BCA ���⵰��Ũ�ȡ�������Ʒ������ 12% ʮ����������ƾ۱�ϩ����������Ӿ(sodium dodecyl sulfate-poly-acrylamide gelelectrophoresis��SDS-PAGE) ��Ӿ��NC תĤ����֬�̷۷������Է�պ��� PCNA(1�� 200)��β-actin(1�� 2 000)��4 ���ҹ��I��Dye 800CW ���ӵĶ���(1�� 10 000)�ܹ���� 1 h���� Odyssey ����ӫ�����ϵͳ��⵰����Ժ������� Image J �����Ե��ױ����������лҶ�ɨ�輰����������
����6) �˿�����ʵ��:������ PASMCs ������ 6 �װ��У������� 100% ��Ϻ��� 1 mL ǹͷ�ڿװ�ײ���2 �� 3 ��ƽ�еĻ��ۣ��� 2 �鴦�� 48 h �ͱ����¹۲�ϸ���˿����������
����1�� 3 ͳ��ѧ����
Ӧ�� Graph Pad Prism 5�� 0 ������ͼ������ SPSS13�� 0 ͳ��ѧ�����������ݴ��������ݲ���ƽ��ֵ ± ����(x�� ± SE)��ʾ�������������Ƚϲ��õ����ط����������������Ƚϲ��� LSD ͳ��ѧ�������� P ��0�� 05Ϊ������ͳ��ѧ���塣
����2 ���
����2�� 1 ϸ����̬ѧ����������ӫ�����
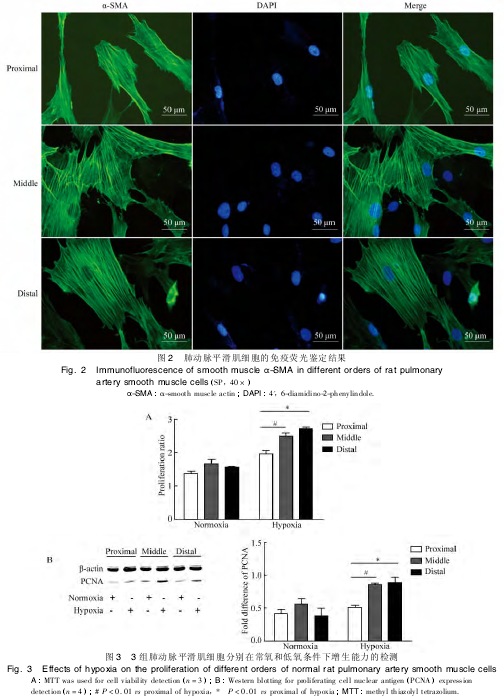
��������������۲죬3 ��ζ�����֯��ԭ������ 2 ��3 d ������ϸ������֯����������ϸ�������������ʳ����η���״��7 �� 10 d ��ϸ�����ںϣ����ֳ����͵�ƽ����ϸ��“�� �� ��”״�����ص�(ͼ1) ���� 3 ��ϸ������̬��δ���ֳ����Բ��졣α-SMA��ϸ������ӫ����ʾ��3 ��ϸ����ϸ�����о��н϶���ϸ��������ƽ�е�˿״��ɫӫ�⣬��Ϊƽ����ϸ�����еļ������ף�������ʵ��������������ϸ����ҪΪ PASMCs(ͼ 2)��
��
��
2�� 2 ������ 3 ����� PASMCs �������ܵIJ�ͬӰ��
�ֱ�� 3 �� PASMCs ���г���(DMEM + 2%FBS��5% CO2��21% O2��37 ��) �͵���(DMEM + 2%FBS��5% CO2��3%O2��37 ��)��Ԥ48 h ���� MTT �����ϸ�����������������£�3 �� PASMCs ϸ�������ֵ�ֱ�Ϊ(1�� 370 ± 0�� 700��1�� 661 ± 0�� 136��1�� 568 ±0�� 144��n = 3��P �� 0�� 05)��������ͳ��ѧ���塣���ڵ���������С�ζ������ж����� PASMCs ϸ�������ֵ�ֱ�Ϊ2��504 ±0�� 093 �� 2�� 729 ±0�� 038�����Ÿ��ڴ�ζ�����PASMCs �������ֵ1��953 ±0��116���������ͳ��ѧ����(n =3��P ��0�� 01��ͼ 3A)��˵����������£���С������Ѫ��ƽ����ϸ�����и�ǿ������������
����PCNA ������ϸ���˿�ԭ����ϸ�� DNA �ϳɹ�ϵ���У���ϸ������������������Ҫ���ã�����ﺬ������ϸ��������������Western blotting �Ľ��Ҳ��ʾ�����ڳ��������£�3 �� PASMCs �� PCNA ���ﺬ���Ҷ�ֵ�ֱ�Ϊ(0�� 415 ± 0�� 052��0�� 551 ± 0�� 089��0�� 381 ±0�� 108��n = 4��P �� 0�� 05)�������Ա�����졣������Ԥ����С�ζ����� PASMCs �� PCNA ���ﺬ���������ߣ��ֱ�Ϊ 0�� 853 ±0�� 023 �� 0�� 885 ±0�� 089�����Ÿ��ڴ�ζ����� PASMCs �� PCNA ���ﺬ��(0�� 509 ±0�� 028)���������ͳ��ѧ����(n = 4��P �� 0�� 01��ͼ3B)����һ��˵����С�ζ���ƽ����ϸ�������������ڴ�ζ���ƽ����ϸ����
����2�� 3 ������ 3 ����� PASMCs Ǩ�ƹ��ܵIJ�ͬӰ��
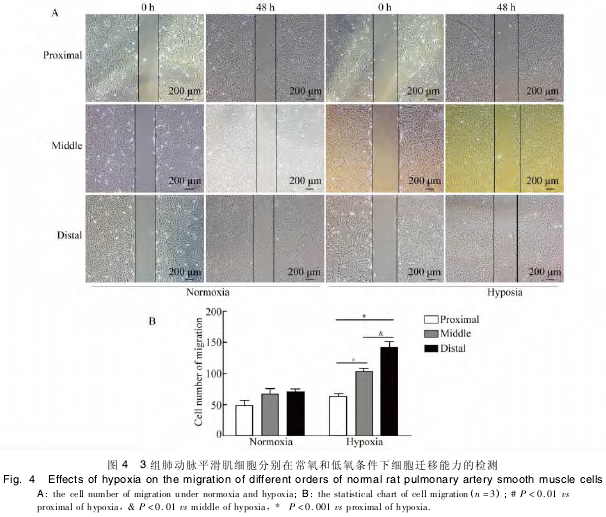
����ѡȡ����״̬���õ�ͬһ�� PASMCs ����ʵ�飬��ϸ�����ں�����Ѫ�� DMEM ���������� 24 h ʹϸ��ͬ�����������ǹͷ��������ײ��� 2 ��3 ��ƽ�еĻ��ۣ����շ���������賣���͵�����Ԥ 48 h �ֱ��Ǩ�Ƶ��հ������ϸ���������м��������������£�3 �� PASMCs ��ϸ��Ǩ�������ֱ�Ϊ(49�� 000 ±7�� 810��68�� 000 ± 7�� 767��71�� 000 ± 4�� 041��n = 3��P �� 0�� 05)���������ͳ��ѧ���塣���ڵ�����Ԥ����С�ζ������ PASMCs Ǩ�Ƶ��հ������ϸ�������ֱ�Ϊ102�� 300 ± 4�� 842 �� 141�� 000 ± 9�� 644�������Ÿ��ڴ�ζ����� PASMCs ��ϸ��Ǩ������ 62�� 670 ±5��044���������ͳ��ѧ����(n =3��P ��0��01��ͼ4)��
����
����3 ����
�����ζ�����ѹ���ɶ��ַ�������������Էζ���������ǿ����С�ζ����ع�Ϊ��Ҫ�����������仯�����յ��·�Ѫ���������������ӡ��ζ���ѹ�����ߵĸ����Լ������ζ������ع���Ҫ����Ϊ PASMCs ��������ʴܱڱ���ھ���խ�ȡ�
�������ٴ��ζ�����ѹ�Ļ����Լ��ζ�����ѹ�Ķ���ģ���У������ַ�Ѫ���ع���Ҫ������ĩ�˵�С���������ҳ���չ�ԣ����յ���ĩ��Ѫ�ܵı�������ʾ�ζ�����ѹ��������Ҫ��λ��Զ�˷ζ��������о���ʾ���������һ����������ֱ��Ϊ 600 ��800 μm������������ֱ֧��Ϊ 400 �� 600 μm������������ֱ֧��Ϊ 100 �� 400μm�����ļ�������֧�����ϣ�����ֱ��ֻ�в���100μm��
�����ڵ���28 d ��ֱ��С��200 μm ��С�ζ����Ѿ�������ʧ���ڵ����Է�Ѫ������ʵ���У�С�����������������Ը��ڴ�����Ѫ��ֱ���Ķ����仯��ʾ��С��500 μm ����С�ζ�����һ�����Եļ��ٹ��̣������������ŵı仯������ֱ��Ϊ 200 �� 300 μm �ķζ����С����Ƕ��ڲ�ͬ�����Ѫ�ܷ�Ӧ�Բ�ͬ��ϸ������ӻ���Ŀǰ�в������
�������о����ý�ԭø������ԭ�������������� 3 ��PASMCs����õ� PASMCs �ʳ����Σ�����2 ��3 d ��ʴ�״��������������7 ��10 d ��ϸ���ںϳ�“��-��”״���������ֳ����͵�Ѫ��ƽ����ϸ����̬ѧ������ƽ����α-SMA ������Ѫ��ƽ����������������ס���ʵ��ͨ������۽�ӫ����������ӫ��Ⱦɫ���֣����� 3�� PASMCs ����ӫ��Ⱦɫ�����Է�Ӧ���ɼ����͵ġ��������е���ɫ��ά��֤ʵ PASMCs ����ƽ���� α-SMA ���ף�����Ѫ��ƽ����ϸ��������ѧ�ṹ������
����PASMCs ����������ϳ���ת��������������ǿ���ӷζ�����ĤǨ������Ĥ(Ǩ�ƹ�����ǿ)�ǵ������·�Ѫ���ع��Ĺؼ����ڡ���ʵ�齫ԭ������ PASMCs �ֱ�¶�ڳ�������������У��� MTT ��Western blotting �ķ������ϸ��������������˿�����ʵ����ϸ����Ǩ�������������ڵ�����������С�ζ����� PASMCs ��������Ǩ�����������Ÿ��ڴ�ζ����� PASMCs����� 3 ��ζ�����ȷ�������������Ѵ���ijЩDZ�ڵIJ��죬ʹ���ڵ���ʱ��С�ζ����������У���ʹ�������Ѫ���ع���
�����������о���Ϊ������ֲ��ڷεĴ�����С������ƽ�������ԵIJ�ͬ���в�ƽ����ϸ���ڷν�����Զ�˶�����ϸ������ϵIJ�ͬ�йء���С����ƽ����ϸ����ϸ�����ͣ�ϸ���ڹǼܵ��ı��������ͨ���Ŀ��������������ͬ�����������ŵIJ��죬���п��������������ܵ�ԭ��֮һ��ȱ������ͨ������ K+ͨ�����ٽ�Ĥ��ȥ�������Ӷ����� L-�� Ca2 +ͨ������ʹ��С�ζ��������������ڵ��������£�ƽ����ϸ���� K+ͨ���ı����빦�ܲ�ͬ��Ӱ�쵰�ױ����ת¼����Ҳ��ͬ��
�������о��д�������㡣����ֻ�Ǹ���Ѫ�ܵķ�֧���ζ����ֳ��� 3 �飬��û���ϸ�ĸ���Ѫ��ֱ�����л���;��Σ����о�ֻ����ϸ��ˮƽ�Ͻ��е���ع��ܵ��о�����û������֯ˮƽ���о���������3 ��Ѫ���ع���Ӱ��;���������ڵ����� 3 �� PASMCs�������ϸ�����ܣ������ϸ�������Ĺ���Ӱ���д���һ�������о���
�����������������о���ϸ��ˮƽ��֤ʵ�˷δ���С�ζ��� PASMCs ȷʵ�����Ź����ϵIJ��죬�ڵ��������£���С�ζ����� PASMCs ������Ǩ��������ǿ����ʵ����Ϊ��һ���о���С���������ع��Ļ����ṩ��һ�����������ݡ�
����4 �����
������1�� Sklepkiewicz P��Sehermuly �� T��Tian X��et al�� Glycogensynthasekinase 3 beta contributes to proliferation of arterialsmooth muscle cells in pulmonary hypertension��J�ݣ� PLoSOne��2011��6(4) :e18883��
��2�� Archer S L��Wu X C��Thebaud B��et al�� Preferential ex-pression and function of voltage-gated�� O2-sensitive K+channels in resistance pulmonary arteries explains regionalheterogeneity in hypoxic pulmonary vasoconstriction: ionicdiversity in smooth muscle cells��J�ݣ� Circ ��es��2004��95(3):308-318��
������3�� Schwenke D O��Pearson J T��Umetani K��et al�� Imaging ofthe pulmonary circulation in the closed-chest rat using syn-chrotron radiation microangiography��J�ݣ� J Appl Physiol(1985)��2007��102(2):787-793��
������4�� McLaughlin V V��McGoon M D�� Pulmonary arterial hyper-tension��J�ݣ� Circulation��2006��114(13):1417-1431��
������5�� Humbert M��Morrell N W��Archer S L��et al�� Cellular andmolecular pathobiology of pulmonary arterial hypertension��J�ݣ� J Am Coll Cardiol��2004��43( Suppl 12):13-24��
������6�� Madden J A��Dawson C A��Harder D �ң� Hypoxia-inducedactivation in small isolated pulmonary arteries from the cat��J�ݣ� J Appl Physiol��1985��59(1):113-118��
������7�� Hislop A����eid L�� New findings in pulmonary arteries ofrats with hypoxia-induced pulmonary hypertension��J�ݣ� Br JExp Pathol��1976��57(5):542-554��
������8�� Leik C E��Willey A��Graham M F��et al�� Isolation and cul-ture of arterial smooth muscle cells from human placenta��J�ݣ� Hypertension��2004��43(4):837-840��
������9�� Barman S A��Zhu S��Han G��et al�� cAMP activates BKCachannels in pulmonary arterial smooth muscle via cGMP-de-pendent protein kinase��J�ݣ� Am J Physiol Lung Cell MolPhysiol��2003��284(6):L1004-1011��
������10�� Suzuki Y J��Day �� M��Tan C C��et al�� Activation of GA-TA-4 by serotonin in pulmonary artery smooth muscle cells��J�ݣ� J Biol Chem��2003��278(19):17525-17531��
������11�� Stiebellehner L��Frid M G����eeves J T��et al�� Bovine distalpulmonary arterial media is composed of a uniform popula-tion of well-differentiated smooth muscle cells with low pro-liferative capabilities��J�ݣ� Am J Physiol Lung Cell MolPhysiol��2003��285(4):L819-828��
������12�� Archer S L�� Diversity of phenotype and function of vascularsmooth muscle cells��J�ݣ� J Lab Clin Med��1996��127(6):524-529��
������13�� Bakhramov A��Evans A M��Kozlowski �� Z�� Differentialeffects of hypoxia on the intracellular Ca2 +concentration ofmyocytes isolated from different regions of the rat pulmonaryarterial tree��J�ݣ� Exp Physiol��1998��83(3) :337-347��
������14�� ë�ã���Ծ�㣬���ܣ��ȣ� �����Ƿ����Ƶ������˷ζ���ƽ����ϸ����ѹ�����Լ�ͨ�����µ����ã�J�ݣ� ��ҽ�ƴ�ѧѧ����2013��33(2):224-231