摘 要: 以芦笋种植土壤为试材, 应用巨大芽孢杆菌 (Bacillus megaterium) 、枯草芽孢杆菌 (Bacillus subtilis) 、凝结芽孢杆菌 (Bacillus coagulans) 和荧光假单孢菌 (Pseudomonas fluorescens) 等比混合的复合菌剂加入蚯蚓粪制备成生物有机肥, 采用完全随机设计, 探究了生物有机肥对土壤酶活性及土壤细菌群落结构的影响。试验设生物有机肥处理4个 (A、B、C、D) 、复合肥处理1个 (E) 、以不施肥为对照 (CK) 。结果表明:处理90d施用生物有机肥对土壤酶活性影响明显, 其中800g·m-2生物有机肥处理土壤脲酶与纤维素酶活性较CK增幅最大, 600g·m-2生物有机肥处理蔗糖酶、过氧化氢酶活性较CK增幅最大, 同时发现PCR-DGGE结果中800g·m-2生物有机肥处理在各阶段细菌多样性表现最好, 其中90dShannon-Wiener指数相对CK和复合肥处理分别增加26.05%、93.87%, 丰富度指数增加24.92%和56.99%。并发现施用生物有机肥后土壤中巨大芽孢杆菌、枯草芽孢杆菌的含量提高, 抑制了有害细菌繁殖。90d测定土壤肥力, 指标显示600g·m-2生物有机肥处理影响土壤全氮、速效磷、速效钾含量最明显, 分别比复合肥处理增加6.34%、37.31%、70.49%;而800g·m-2生物有机肥处理影响土壤全磷、全钾、有机质含量最明显, 分别比复合肥提高了1.43%、5.82%、18.82%。综上所述, 施用生物有机肥能增加功能性有益细菌的含量, 明显提高土壤细菌群落多样性与丰富度, 从而提升土壤酶活性和土壤肥力。
关键词: 生物有机肥; 细菌多样性; 酶活性; 土壤肥力;
Abstract: Asparagus planting soil was the test material and the experiment was a completely randomized controlled study.We mixed 4 strains (Bacillus megaterium, Bacillus subtilis, Bacillus coagulans and Pseudomonas fluorescens) and wormcast to manufacture bio-organic fertilizer.The study investigated the effects of different bio-organic fertilization amount on soil fertility factors and soil bacterial community structure.The experiment consisted of 4 bio-organic fertilizer treatments (A, B, C, D) , 1 compound fertilizer (E) , and no fertilization (CK) .The results showed that the enzyme activities of urease and cellulase in the soil treated with 800 g·m-2 bio-organic fertilizer treatment for90 days increased the most, the activity of sucrase and catalase increased the most in the 600 g·m-2 bio-organic fertilizer treatment and 800 g·m-2 bio-organic fertilizer treatment had the best bacterial diversity in all stages through the PCR-DGGE results.The Shannon-Wiener index increased by26.05%and 93.87% with respect to CK and compound fertilizer treatment at 90 days, respectively.And the Margalef′s index increased by 24.92% and 56.99%, respectively.The Bacillus megaterium and Bacillus subtilis content increased and the harmful bacteria decreased after used bio-organic fertilizer.The soil fertility indicators showed that the 600 g·m-2 bio-organic fertilizer treatment significantly affected soil total nitrogen, available phosphorus and available potassium, which were6.34%, 37.31%and 70.49%higher than those of the compound fertilizer treatment, respectively.And the 800 g·m-2 treatment significantly affected soil total phosphorus, total potassium and organic matter, which were 1.43%, 5.82% and 18.82% higher than those of the compound fertilizer treatment, respectively.Bio-organic fertilizer could increase the content of functionally beneficial bacteria, increase the diversity and richness of soil bacterial community, thus enhance soil enzyme activity and soil fertility.
Keyword: bio-organic fertilizer; bacterial diversity; enzyme activity; soil fertility;
芦笋 (Asparagus officinalis) 属百合科天门冬属多年生植物, 又名石刁柏。不仅是我国, 还是世界各地人们喜爱的蔬菜[1]。我国是目前世界第一大芦笋生产和出口国[2]。多年来由于种植户偏施化肥造成土壤板结[3]。传统有机肥养分含量低, 用量大, 推广起来容易加剧土传病害和面源污染, 因此开发新型生物有机肥具有重要意义。生物有机肥是将腐熟有机肥和具有特定功能的微生物复合从而形成的一种新型肥料, 兼具微生物菌肥和有机肥的优点[4]。能够显着提高土壤有机质含量, 增强土壤中微生物活性[5];分解土壤中难溶的磷、钾化合物, 减少养分的淋溶与农业面源污染, 也是实现农业可持续发展、改善农业生态环境和发展循环经济的有效途径。

当前, 生物有机肥的效应研究主要在大田作物[6]、经济作物[7], 或经济价值高的植物如蓝莓[8]上。而施用功能性的生物有机肥对芦笋种植土壤肥力的影响和对其根际环境, 特别是细菌群落结构的影响尚鲜见报道。该研究以芦笋种植土壤为试验材料, 探究施用生物有机肥后土壤肥力因子的变化, 并采用PCR-DGGE的方法, 研究生物有机肥对芦笋种植土壤细菌群落多样性的影响, 为进一步系统开展芦笋根际土壤微生物群落结构形成规律和提升生物有机肥的质量品质提供参考基础。
1、 材料与方法
1.1、 试验地概况
试验于2017年4—8月在四川师范大学成龙校区科研基地温室中进行。种植土壤为山地黄壤黏土, 土壤有机质含量23.38g·kg-1, 全氮含量0.74g·kg-1, 全磷含量0.86g·kg-1, 全钾含量6.68g·kg-1, 速效磷含量417.11mg·kg-1, 速效钾含量410.52mg·kg-1。
1.2、 试验材料
供试芦笋苗选用格兰德杂交芦笋种子 (F1) 播种的2年生实生苗。生物有机肥是以蚯蚓粪为基质, 加入实验室已筛选出来的巨大芽孢杆菌 (Bacillus megaterium) 、枯草芽孢杆菌 (Bacillus subtilis) 、凝结芽孢杆菌 (Bacillus coagulans) 和荧光假单孢菌 (Pseudomonas fluorescens) 组成的菌系。各菌种分别培育在50mL种子培养基中生长24h后将其等比例混合, 混后菌液加入到玉米干粉配制的发酵液中, 使用液体发酵罐 (5L, 镇江东方生物工程设备技术有限责任公司GBJS-5L) 混合发酵3d, 3d后 (使用浓度梯度稀释涂布法测得) 发酵液中有效活菌数为0.7亿cfu·mL-1, 再将发酵液与蚯蚓粪以0.3∶1 (体积比) 的比例混匀获得试验用生物有机肥。生物有机肥技术指标为有机质≥45%, N+P5O2+K2O≥5%。
1.3 、试验方法
试验设6个处理。A:200g·m-2生物有机肥处理;B:400g·m-2生物有机肥处理;C:600g·m-2生物有机肥处理;D:800g·m-2生物有机肥处理;E:复合肥处理;CK:不施肥为对照 (注:各处理的名称分别以A、B、C、D、E、CK表示, 下同) 。试验采用单因素完全随机试验, 每处理3次重复。
分别于30、60、90d采集土样。样品为表层土壤下0~20cm处, 每处土样S形采集5钻土, 充分均匀后作为一个样剔除根系、石块等杂质, 用密封袋分成2份封样带回实验室;1份自然风干, 研磨土样过60目筛子, 常温放置, 储存待用, 进行土壤肥力指标测定[8]。另1份用于提取土壤总DNA。
1.4 、项目测定
1.4.1、 土壤肥力因子测定
土壤总氮含量测定采用半微量凯氏定氮法;土壤总磷含量测定采用钼锑抗显色分光光度法;土壤全钾含量测定采用火焰分光光度法;土壤有机质含量测定采用重铬酸钾容量法;土壤速效钾含量测定采用醋酸铵提取-火焰光度法;土壤速效磷含量测定采用碳酸氢钠浸提-钼锑抗显色分光光度法[9]。
1.4.2 、土壤酶活测定
脲酶采用靛酚蓝比色法测定, 以单位时间内1g风干土催化生成NH3-N的mg表示;蔗糖酶采用DNS比色法测定, 以单位时间内1g风干土催化生成葡萄糖的mg表示;纤维素酶采用DNS比色法测定, 以单位时间内1g风干土催化生成葡萄糖的mg表示;过氧化氢酶采用硫酸钛分光光度法, 以单位时间内1g风干土中水解的H2O2μg数表示[10]。
1.4.3、 土壤微生物总DNA提取及PCR扩增
采用康为世纪Soil Genomic DNA Kit试剂盒提取土壤样品总DNA, 在0.8%的琼脂糖凝胶电泳检测后保存在-20℃。总DNA样品PCR扩增在Bio-Rad的基因扩增仪 (S1000 Thermal Cycler美国) 上进行。PCR反应体系:扩增引物为细菌通用引物27F与1942R, 其序列分别为27F:5′-AGAGTTTGATCCTGGCTCAG-3′;1492R:5′- (GC) -TACGGCTACCTTACGACTT-3′;GC结构碱基为:5′-CGCCCGCCGCGCGCGGCGGG-CGGGGCGGGGGC-ACGGGGGG-3′, 上、下游引物 (10μmol·L-1) 各1μL;3μL DNA模板;25μL 2×Taq PCR MasterMix (天根) ;ddH2O补足至50μL。PCR反应条件:采用Touchdown程序, 94℃预变性5min;94℃变性30s, 68℃退火30s, 每个循环降低1℃, 72℃延伸1 min, 10个循环;94℃变性30s, 58℃退火30s, 72℃延伸1min, 20个循环;72℃延伸10min。
1.4.4、 DGGE条件和分析
采用变性凝胶梯度电泳 (DGGE) 。电泳条件:8%聚丙烯酰胺凝胶, 变性梯度为30%~60%, 90V恒定电压下60℃下电泳9h[11]。
1.4.5、 凝胶指纹图谱的生物信息学分析
DGGE凝胶指纹图谱条带用Quantity One软件分析, 测得条带的相对亮度值为该菌种的相对多度 (Pi) 。用UPGMA法进行聚类分析, 并采用香浓指数 (Shannon-Wiener index) 、均匀度指数 (Pielou′s index) 、丰富度指数 (Margalef′s index) 计算土壤样品细菌群落的多样性[12]。
1.4.6、 聚丙烯酰胺凝胶条带测定
在凝胶能成像系统 (Bio-Rad/Ge1DoctmXR+) 成像之后进行切胶, 切下的条带使用OMEGA公司Poly-Gel DNA Extraction Kit试剂盒回收聚丙烯酰胺凝胶条带。再使用27F与1942R细菌上下游通用引物经PCR扩增送鉴定。
1.5、 数据分析
采用Excel 2019软件对数据进行处理, 采用SPSS 18.0软件对数据进行单因素方差分析 (P<0.05) , 采用Origin 2017软件进行图片绘制。
2 、结果与分析
2.1、 不同施肥处理对土壤酶活性的影响
不同施肥处理对土壤酶活性随时间的影响如图1所示, 不同处理对土壤不同酶活性有不同的影响。土壤脲酶活性如图1a所示, D处理活性最高, 而E处理与CK随处理时间的增加脲酶活性降低, 且E处理酶活性降低速率更快。土壤蔗糖酶活性如图1b所示, E处理蔗糖酶活性下降明显, 而空白处理呈缓慢的下降趋势, B、C、D处理随时间的增加其增长速率无明显的差异, 60d以后A处理蔗糖酶活性有所下降。土壤纤维素酶活性如图1c所示, 使用E处理的土壤纤维素酶活性随处理时间的增加呈缓慢的下降趋势, CK土壤纤维素酶活性随时间的增加而缓慢增加, A、B、C、D处理土壤各组土壤纤维素酶活性均明显的增加, 且D处理土壤纤维素酶活性增加约明显。土壤过氧化氢酶活性如图1d所示, E处理过氧化氢酶活性下降明显, 而CK土壤过氧化氢酶活性保持稳定, A、B处理随处理时间的增加土壤过氧化氢酶活性呈持续的增加, C、D处理随处理时间的增加在60d之前土壤过氧化氢酶活性增加明显, 处理了60d之后, 土壤过氧化氢酶活性降低, 且D处理过氧化氢酶活性下降大于C处理。
图1 不同处理对土壤脲酶活性 (a) 、蔗糖酶活性 (b) 、纤维素酶活性 (c) 、过氧化氢酶活性 (d) 的影响
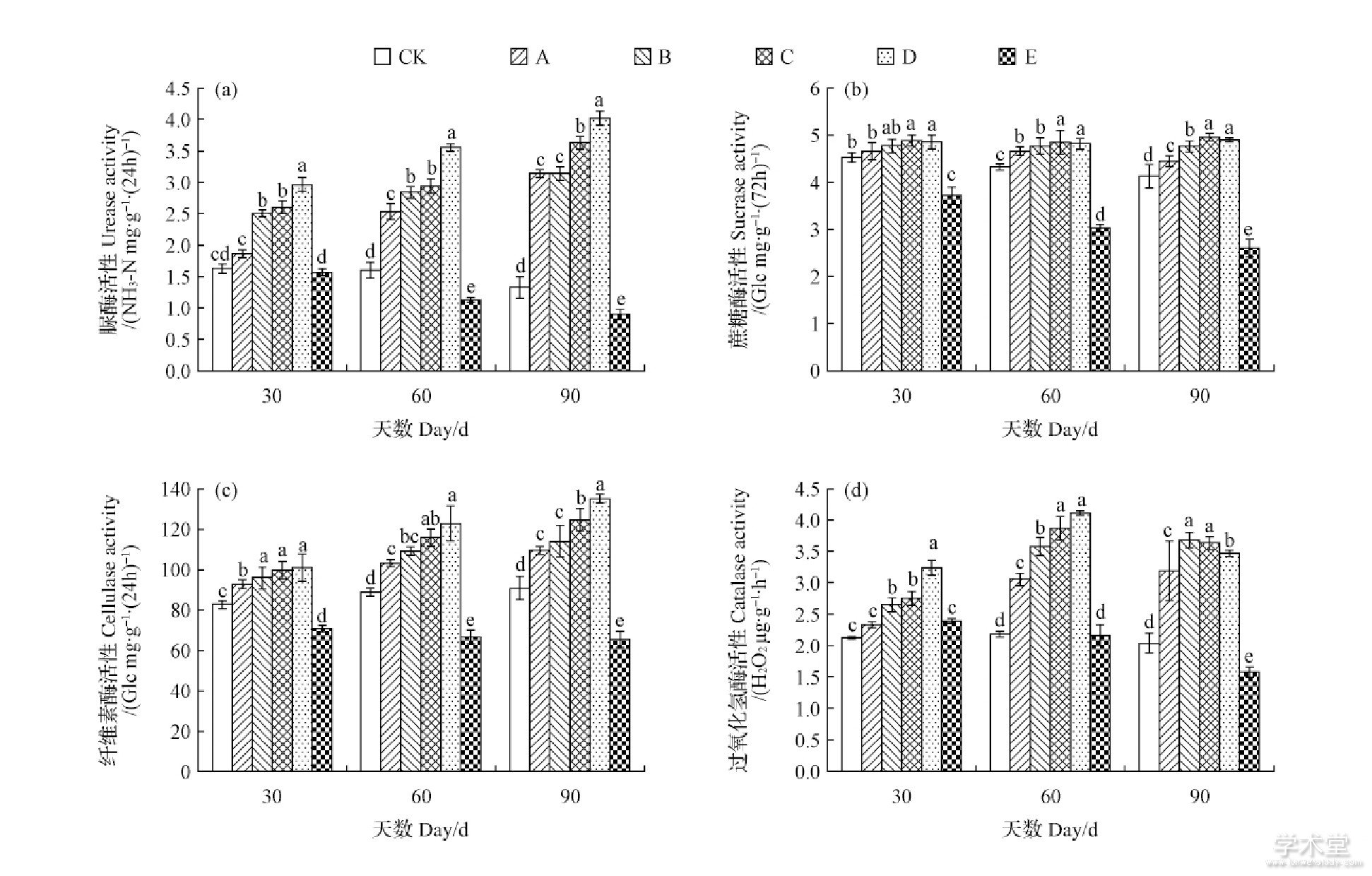
Fig.1 Effects of different treatments on activities of soil urease (a) , sucrase (b) , cellulase (c) , catalase (d)
细菌PCR产物的DGGE图谱可以直观的反应各处理对应的迁移信息和条带的数目, 运用Quantity One软件对图谱进行基本的背景排除然后通过泳道、条带识别匹配等步骤可得到泳道间的对比分析结果图 (图2~4) 。该研究根据电泳图谱中每条条带的信息对土样中细菌的Shannon-Wiener (H’) 、均匀度指数 (J) 、丰富度指数 (S) 这3类指标进行了综合性分析, 结果如表1所示。各处理中Shannon-Wiener指数差异显着, 在施肥后60d时, D处理Shannon-Wiener指数最高为3.389, 丰富度也最大为6.913。复合肥处理Shannon-Wiener指数最低为1.468, 丰富度也最低为3.257。随着生物有机肥施用量增加和处理时间的增长, 土壤中的细菌Shannon-Wiener指数与丰富度指数均呈先增大后减少的趋势, 均匀度指数无显着变化, D处理Shannon-Wiener指数和丰富度指数分别在30、60d时为最大值, 而在处理90d时C处理Shannon-Wiener指数最大, B处理生物有机肥组丰富度指数最大。
表1 不同施肥处理后30、60、90d根际土壤细菌遗传多样性分析
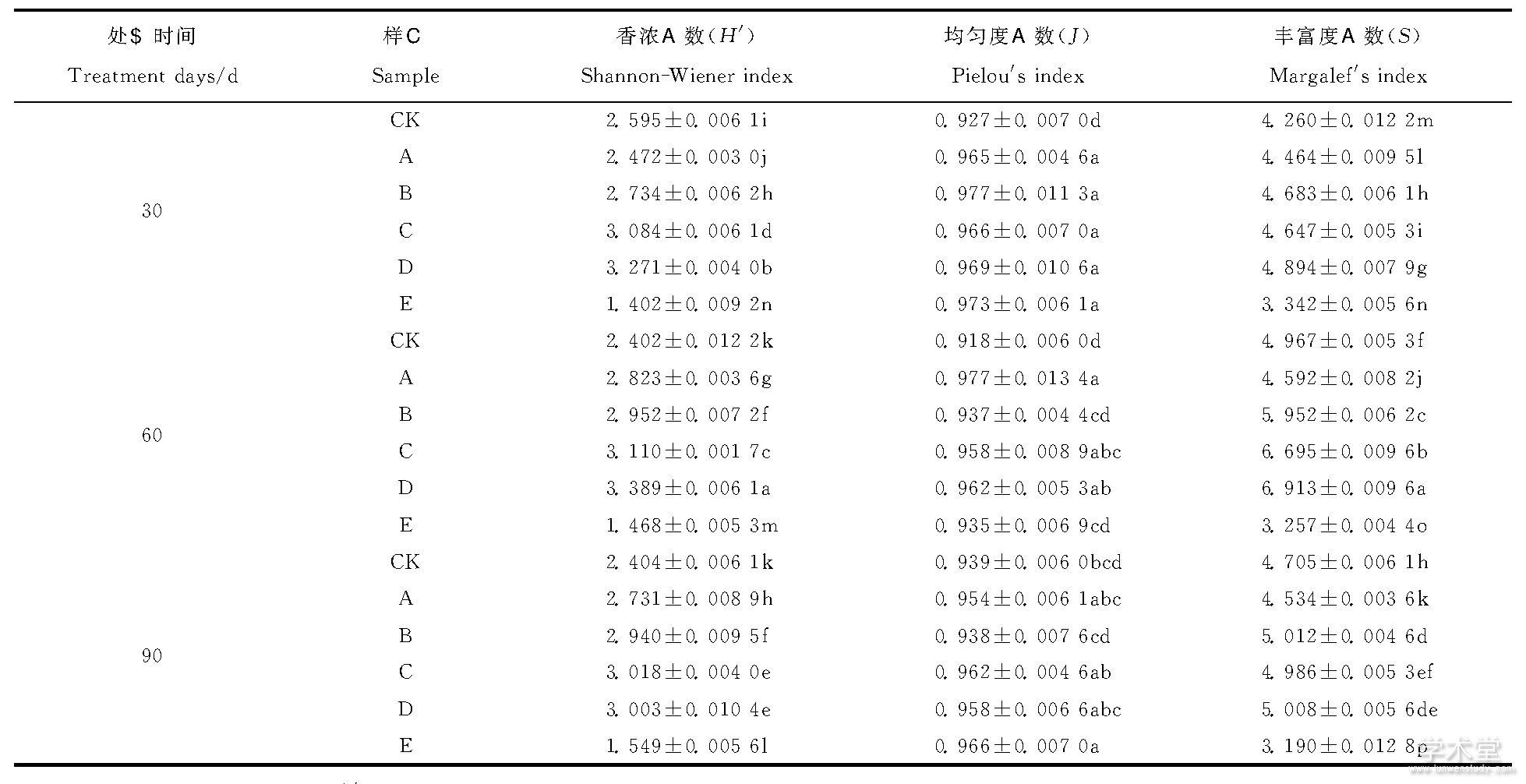
注:同列不同小写字母表示5%的差异水平, 下同。
Note:The list of different lowercase letters with 5%difference in level, the same below.
经过切胶、回收、测序发现图中条带a、b、c、e分别为试验初期筛选的巨大芽孢杆菌、枯草芽孢杆菌、凝结芽孢杆菌和荧光假单芽孢菌。条带d、f为在土壤中测序后发现的大肠杆菌与亚硝化单胞菌。a、b、c、e 4类菌随着生物有机肥使用量增加, 条带明显变亮即此4类菌在土壤中的含量逐步增加。条带b在B、C、D处理的条带明暗程度随处理浓度的增加逐渐变亮, 而在A、CK处理的2条泳道上明暗程度变化不明显, 说明b条带对应的细菌对人工施加生物有机肥有一定的相关性。而c与e条带则一直在A、B、C、D、CK处理的5个条带上均较为明亮, 说明人为施加生物有机肥对其影响不大。条带d (大肠杆菌) 在A、B、C、D处理的条带均较浅, 而在CK的条带d明显较亮, 说明在施加生物有机肥后对条带d的微生物有一定的抑制作用。
图2 施肥30d土壤16SrDNA V3区DGGE扩增片段 (左) 及电泳比较图 (右)
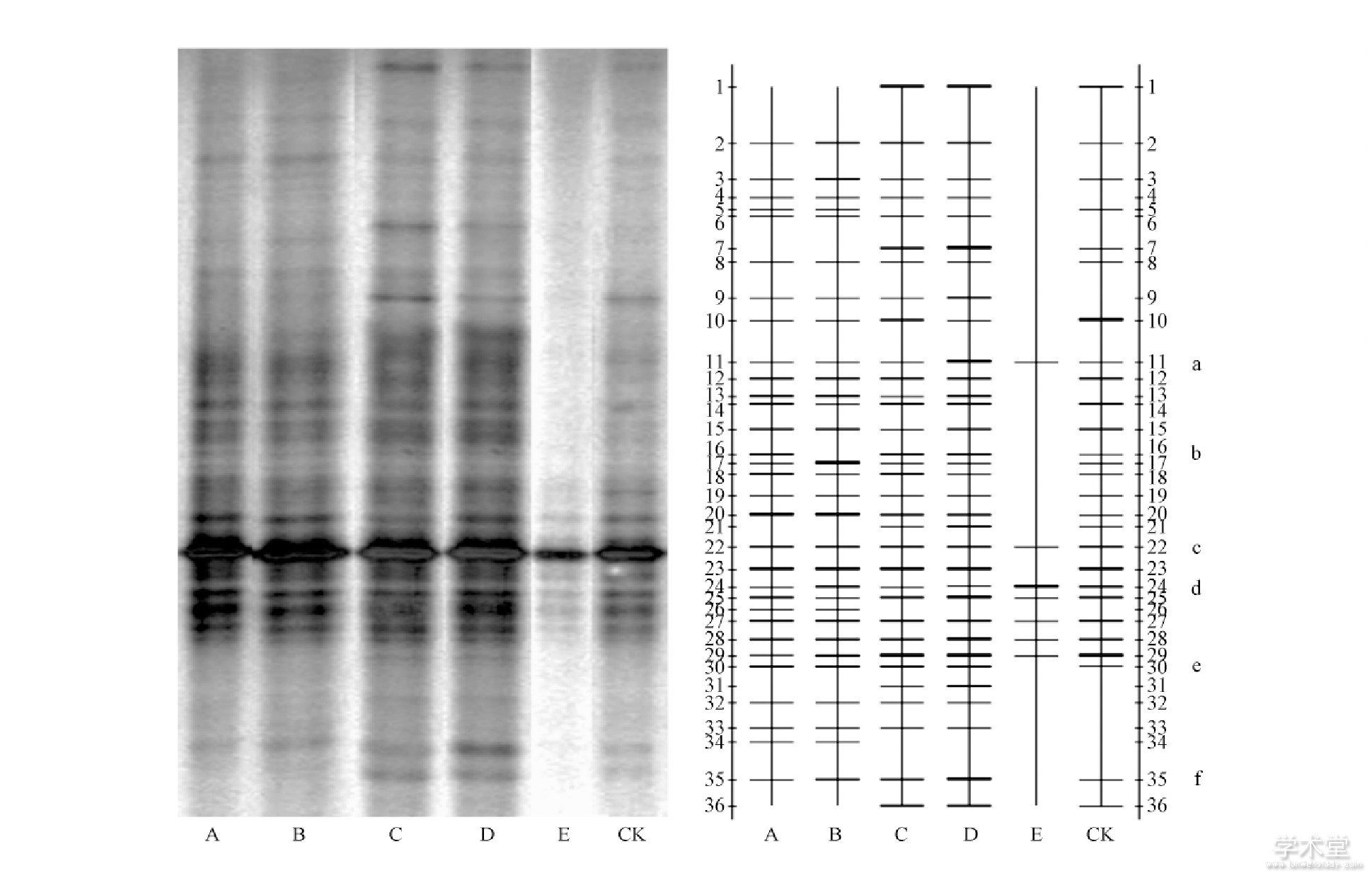
Fig.2 Fertilized soil for 30days the 16SrDNA V3region DGGE amplification fragment (left) and land compare (right)
图3 施肥60d土壤16SrDNA V3区DGGE扩增片段 (左) 及电泳比较图 (右)
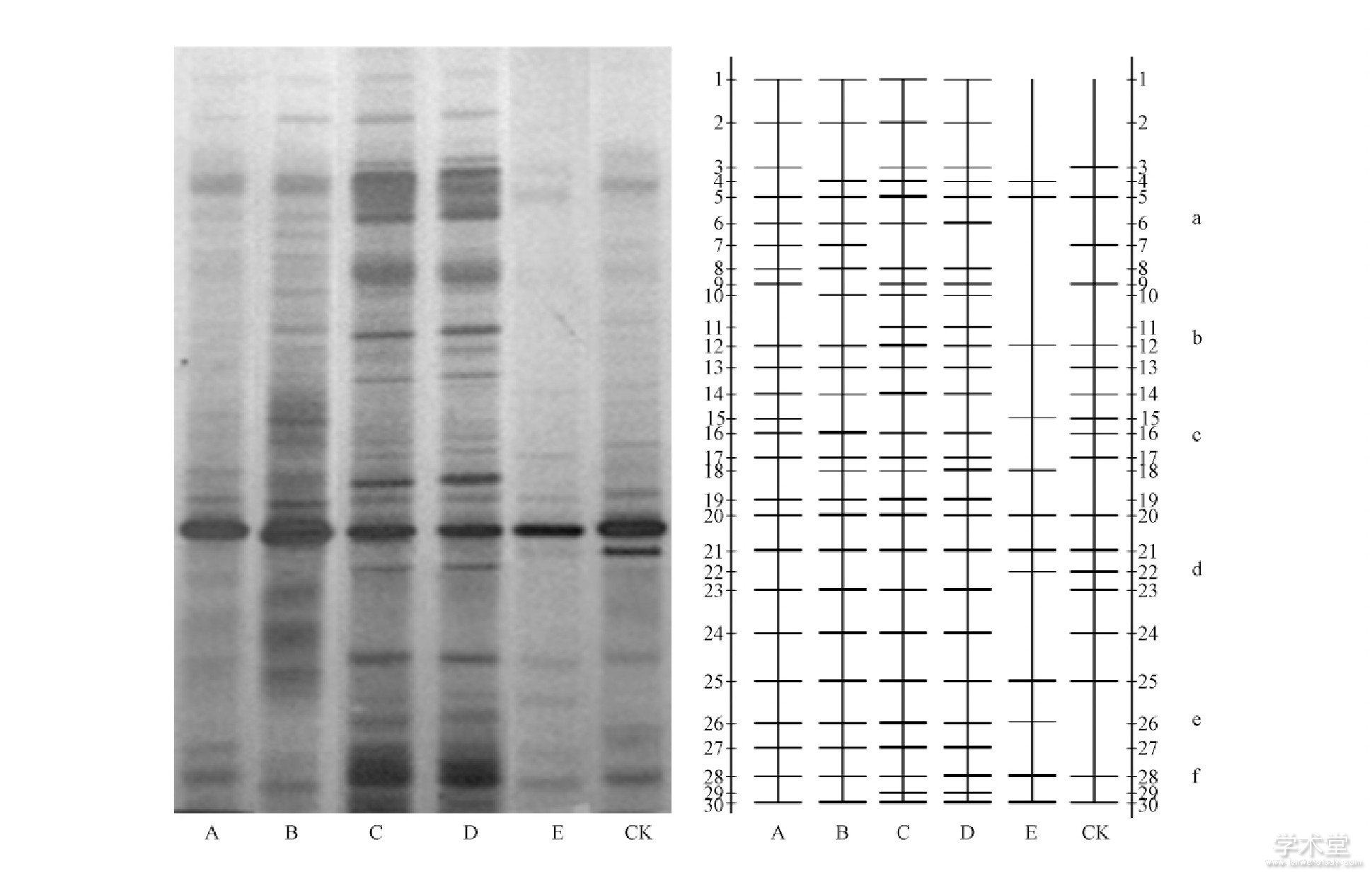
Fig.3 Fertilized soil for 60days the 16SrDNA V3region DGGE amplification fragment (left) and land compare (right)
图4 施肥90d土壤16SrDNA V3区DGGE扩增片段 (左) 及电泳比较图 (右)

Fig.4 Fertilized soil for 90days the 16SrDNA V3region DGGE amplification fragment (left) and land compare (right)
利用相似性矩阵, 通过UPGMA对不同细菌群落结构相似性进行聚类分析, 结果如图5所示。复合肥组与其它处理相似结构较低, A、B处理聚为一类, C、D处理聚为一类, E处理为一类, CK为一类。比对DGGE图谱分析, A、B、C、D处理与E处理对土壤微生物量均有影响, A、B、C、D处理对维持和改善根际土壤细菌多样性效果明显, E处理对土壤细菌多样性有减少效应。
图5 施肥90d土壤UPGMA群落结构相似性的聚类分析
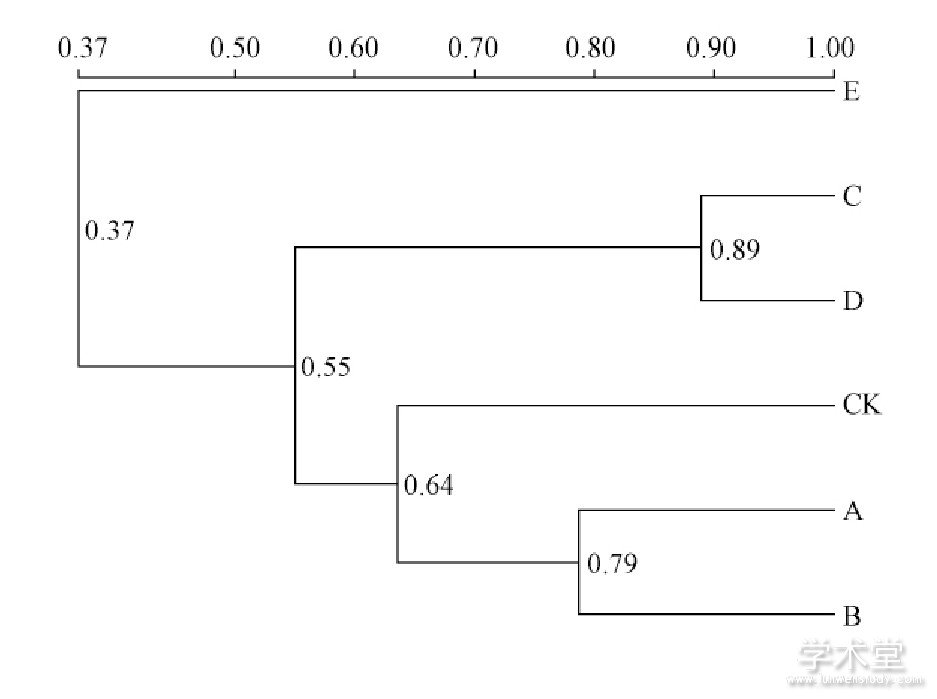
Fig.5 Clustering analysis of similarity of soil UPGMA community structure by fertilized for 90days
2.2、 生物有机肥对土壤肥力主要因子的影响
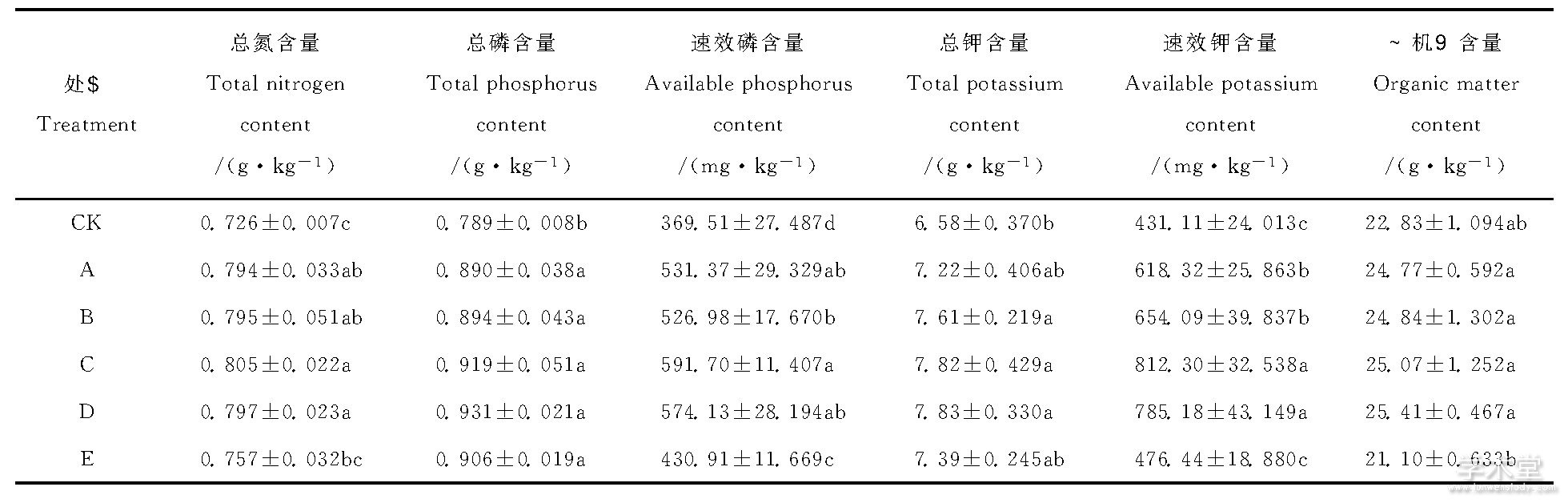
从表2可以看出, 不同处理对土壤总氮含量的影响效果不同, E处理比CK提高4.3%, 而生物有机肥根据不同施肥浓度总氮提高9.4%~10.8%;不同处理间总磷含量并无显着差异, 而各处理间速效磷含量有显着差异, E处理与其它处理差异显着 (P<0.05, 下同) , 速效磷含量最高的C处理相对CK增加了60.1%;D处理总钾含量最高, 其次为C处理分别相对CK提高了19.0%与18.8%, A、B、C、D处理与E处理相比无显着差异, 而各处理间速效钾含量同样有所差异, A、B、C、D处理速效钾含量均显着高于CK与E处理, 其中C处理含量最高相对于CK速效钾含量提高了88.4%, CK与E处理无显着差异;A、B、C、D处理有机质含量分别与CK差异显着, 其中D处理含量最高, 比CK增加11.3%, E处理与CK相比, 有机质含量降低7.6%。
表2 不同施肥处理90d后土壤肥力主要因子指标值
3、 讨论
3.1、 生物有机肥对土壤酶活性的影响
试验中E处理所测酶活性均随处理时间的活性降低, 这可能是因为施加复合肥使土壤团粒结构遭到了破坏, 而黏粒、小微粒在浇水后随水流缓处沉淀。王利民等[13]研究发现, 土壤蛋白酶、脲酶活性与土壤砂粒含量呈显着负相关, 而与粉粒、粘粒含量均呈显着正相关。结果显示施加生物有机肥提高了土壤脲酶、纤维素酶与过氧化氢酶活性, 可能是施加生物有机肥加快了土壤微生物的繁殖速度, 加上试验所用的生物有机肥中本身含有大量的功能微生物所致。该研究从春季开始到夏季结束, 气温不断升高, 环境因素也是提高土壤酶活性重要原因。樊军等[14]通过对不同施肥条件下冬小麦旱地土壤酶动态参数的测定, 发现有机肥对脲酶影响最大。施用生物有机肥可促进作物根系代谢, 加快微生物繁殖速度, 从而提高土壤酶活性[15]。徐华勤等[16]发现水分和温度对土壤酶活性的影响夏季高于秋季。这也证明了土壤中微生物与环境因素均能对土壤酶活性产生影响。由于不同含量有机质、C/N等影响土壤不同酶活性均有差异, 一般来说, 有机质中C/N含量越低, 微生物活性越高, 土壤酶活性越高。从某种意义上说, 施用有机肥是一种添加酶的措施[16]。
3.2 、生物有机肥对芦笋种植土壤细菌多样性的影响
土壤的品质不仅仅由土壤理化性质决定, 而且与土壤生物学性质息息相关。土壤中微生物的多样性和数量是表征土壤肥力重要的生物学指标。胡可等[5]研究了生物有机肥对土壤微生物活性的影响, 结果表明生物有机肥基料多为有机物, 可丰富土壤中微生物可利用营养, 增加菌群数。这也解释了该试验中添加生物有机肥后土壤微生物多样性与丰富度显着增加, 且施肥量大的处理多样性与丰富度越高的原因。MARSCHNER等[17]研究发现土壤有机质含量越高, 土壤对微生物的缓冲能力就越强, 土壤生物量就越高。该研究同样也得到了这个结论, 施加生物有机肥提高了土壤有机质含量的同时土壤微生物含量也显着提高。SUN等[18]研究了施用有机肥和化肥对土壤细菌群落结构的影响, 结果显示施用了氮磷钾化肥处理的DGGE条带数明显地少于有机肥处理, 化肥处理显着的降低了土壤细菌群落的多样性。该试验对芦笋种植土壤的研究也得出了同样的结果, 施用生物有机肥能提高土壤微生物量, 增强土壤生物活性, 增加土壤中有益菌的含量, 有益菌在生长繁殖过程中产生大量的代谢产物, 促使有机物的分解转化, 能直接或间接为作物提供多种营养和刺激性物质, 促进和调控作物生长。
3.3 、生物有机肥对芦笋种植土壤肥力的影响
土壤中的氮、磷、钾含量是衡量肥力水平的重要指标, 已有研究表明, 在一定含量范围内, 随着土壤氮、磷、钾含量的增加, 生物量和有机质的含量也随之提高[19]。该试验的生物有机肥, 施用到芦笋种植土壤能增加土壤总氮、磷、钾含量, 并显着的提高了土壤有机质含量。KUMAR等[20]研究发现, 生物肥料的施用, 使得土壤团粒结构有机添加物和水稳性团聚体显着影响, 土壤速效氮、磷、钾、硫含量最高。在该试验中施加生物有机肥后, 土壤速效磷、速效钾同样证明了施用生物有机肥对土壤速效磷、速效钾含量有显着的提升。土壤中速效磷、速效钾可表征土壤的供磷、供钾状况, 也是诊断当季土壤有效肥力的指标之一[21], 速效磷、速效钾的含量一般随土壤黏粒含量的增加而减少, 土壤粉粒含量增加而增加。由此推断, 施用生物有机肥, 一方面固氮、解磷、解钾功能的微生物菌系增大了土壤中氮、磷、钾含量, 另一方面生物有机肥颗粒蓬松表面积大, 降低土壤黏粒含量并增大粉粒含量, 从而提高土壤的速效磷、速效钾含量。
4 、结论
该研究发现, 添加功能微生物菌群的蚯蚓粪所制成的生物有机肥中含有益菌群, 对土壤土着微生物有明显活化作用, 而生物有机肥中的有机质可以给土壤中微生物提供更多的营养物质, 从而增加土壤细菌的多样性。PCR-DGGE图谱表明生物有机肥提高了土壤中功能性有益细菌含量, 并能抑制部分有害细菌的生长, 说明施用生物有机肥后土壤根际环境得到改善, 提高了有益菌在土壤中的竞争力。因为土壤微生物环境的改善, 土壤有机质含量显着增加, 从而提高土壤保肥的能力, 所以施用生物有机肥后能够使土壤供肥能力的持久性和稳定性提升, 加上施肥后土壤营养物质显着提高, 添加进生物有机肥中的功能微生物能有效的促进土壤难溶物的分解, 显着提高了土壤的肥力。一定程度上促进了土壤酶活性与作物根系代谢, 增加根系分泌, 加快土壤微生物繁殖速度。施加进土壤的微生物有机肥中的功能微生物自身可能会产生一部分功能酶, 从而能进一步的提高土壤酶的活性。
(该研究受四川师范大学实验技术项目 (SYJS2016007) 资助, 特此致谢。)
参考文献
[1]陈河龙, 马振川, 杨克军, 等.芦笋种质资源营养品质分析及评价[J].热带作物学报, 2018 (6) :1061-1066.
[2]彭柳林, 余艳锋, 周开洪.我国芦笋产业布局优化分析[J].中国农业资源与区划, 2015, 36 (1) :123-127.
[3]李绪良, 宋绍明, 朱占林.川西北地区绿芦笋生长异常及应对措施[J].中国蔬菜, 2015, 1 (9) :87.
[4]LARKIN R P, GRIFFIN T S.Control of soilborne potato diseases using Brassica green manures[J].Crop Protection, 2007, 26 (7) :1067-1077.
[5]胡可, 李华兴, 卢维盛, 等.生物有机肥对土壤微生物活性的影响[J].中国生态农业学报, 2010, 18 (2) :303-306.
[6]DAS S, ADHYA T K.Effect of combine application of organic manure and inorganic fertilizer on methane and nitrous oxide emissions from a tropical flooded soil planted to rice[J].Geoderma, 2014, 213 (1) :185-192.
[7]RASHMI K, SHNAKAR M A, NARAYANASWAMY TK, et al.Role of introduced organics and biofertilizers on soil fertility status and microflora of S36 mulberry garden[J].Mysore Journal of Agricultural Sciences, 2009, 43 (4) :646-649.
[8]孙运杰, 马海林, 刘方春, 等.生物肥对蓝莓根际土壤微生物学特性及土壤肥力的影响[J].水土保持学报, 2015, 29 (3) :167-171.
[9]叶协锋, 杨超, 李正, 等.绿肥对植烟土壤酶活性及土壤肥力的影响[J].植物营养与肥料学报, 2013, 19 (2) :445-454.
[10]关松萌.土壤酶及其研究法[M].北京:农业出版社, 1986:274-339.
[11]王亚男, 李睿玉, 朱晓换, 等.土荆芥挥发油化感胁迫对土壤胞外酶活性和微生物多样性的影响[J].生态学报, 2017 (13) :4318-4326.
[12]陈法霖, 张凯, 郑华, 等.PCR-DGG技术解析针叶和阔叶凋落物混合分解对土壤微生物群落结构的影响[J].应用与环境生物学报, 2011, 17 (2) :145-150.
[13]王利民, 李卫华, 范平, 等.长期培肥下红黄壤区茶园土壤酶活性的变化[J].茶叶科学, 2012, 32 (4) :347-352.
[14]樊军, 郝明德.旱地农田土壤脲酶与碱性磷酸酶动力学特征[J].干旱地区农业研究, 2002, 20 (1) :35-37.
[15]史吉平, 张夫道, 林葆.长期施肥对土壤有机质及生物学特性的影响[J].中国土壤与肥料, 1998 (3) :7-11.
[16]徐华勤, 肖润林, 向佐湘, 等.稻草覆盖、间作三叶草茶园土壤酶活性与养分的关系[J].生态学杂志, 2009, 28 (8) :1537-1543.
[17]MARSCHNER P, KANDELER E, MARSCHNER B.Structure and function of the soil microbial community in a longterm fertilizer experiment[J].Soil Biology&Biochemistry, 2003, 35 (3) :453-461.
[18]SUN H Y, DENG S P, RUAN W R.Bacterial community structure and diversity in a century-old manure-treated agroecosystem[J].Applied and Environmental Microbiology, 2004, 70 (10) :5868-5874.
[19]KANCHIKERIMATH M, SINGH D.Soil organic matter and biological properties after 26years of maize-wheat-cowpea cropping as affected by manure and fertilization in a Cambisol in semiarid region of India[J].Agriculture Ecosystems and Environment, 2001, 86 (2) :155-162.
[20]KUMAR V C, GAJANANA G N, BASAVARAJ B.Influence of sources of organic manures on availability of micronutrients to finger millet (Eleusine coracana, Gaertn) in Alfisal[J].Mapana-Journal of Sciences, 2004 (2) :8-13.
[21]LIANG B, ZHAO W, YANG X, et al.Fate of nitrogen-15as influenced by soil and nutrient management history in a 19-year wheat-maize experiment[J].Field Crops Research, 2013, 144:126-134.





