
精液冷冻保存对动物种质资源的保存具有重要意义,然而与其他哺乳动物相比,猪精子在冷冻过程中更易受到损伤,其冷冻保存效果仍不理想.猪精子在温度低于15 ℃时十分容易受到低温打击(coolshock),表现为质膜完整性受损.质膜损伤往往导致细胞内容物的外泄而使精子死亡,顶体膜损伤会引发顶体酶的泄漏(假顶体反应)而使精子失去受精能力.而冷冻过程中精子质膜的损伤可能与活性氧(Re-active oxygen species,ROS),如过氧化氢(H2O2)、超氧阴离子(O-2·)、羟自由基(OH·)、一氧化氮(NO)以及丙二醛(MDA)等水平的上升有关.
猪精液冷冻保存的降温过程主要包括3个阶段,即从体温(约38.5 ℃)降温至15 ℃进行预平衡(减少精子对低温打击的敏感性),从15℃降温至5℃平衡(使冷冻液中甘油扩散均匀)以及从5℃降温至液氮温度(-196℃).
在不同阶段,精子所受到的损伤也有差异,而这种差异主要与精子细胞内ROS的水平相关.Drobnis等的研究表明,从15℃降温至5℃的过程中,随着胞内ROS水平的上升,精子发生的损伤也逐渐上升,他们认为该阶段也是精子受损的主要阶段.Kim等和Ball等研究发现,液氮冷冻过程会使精子中ROS水平大幅度升高,大量的ROS会氧化细胞膜脂质中的不饱和脂肪酸,使之发生脂质过氧化(Lipid peroxidation,LPO)而损伤精子细胞膜.
虽然很多研究结果都表明,精子细胞膜损伤程度与ROS水平相关,但是精子冷冻过程中ROS水平上升的原因仍不清楚.
目前的研究发现,哺乳动物精子中含有能够清除ROS的酶促防御系统,可以保护精子免受氧化损伤,如超氧化物歧化酶(Superoxide Dismutase,SOD)、过氧化氢酶(Catalase,CAT)和谷胱甘肽过氧化物酶(Glutathione peroxidase,GPX)等.
猪精子冷冻过程中ROS水平的升高是否与抗氧化酶活性的变化有关?精子细胞膜是否因为抗氧化酶活性的变化而发生损伤?针对上述问题,本文主要分析了巴马香猪精液冷冻保存过程中不同降温阶段(鲜精、15 ℃精液、5 ℃精液、冷冻-解冻后精液)精子质膜和顶体完整性、抗氧化酶活性以及ROS水平的变化,从而研究抗氧化酶活性变化对巴马香猪精子质膜完整性的影响.
1 材料和方法
1.1 主要仪器和设备
伟力彩色精子质量检测系统(WLJY-9000型,北京伟力新世纪发展有限公司)、超低温温度计(600-1010K型,美国Barnant公司)、自动高速离心机(Heraeus,德国)、荧光显微镜(Nikon E600,日本)、电热恒温水浴锅、pH测试仪、液氮罐、冰箱、恒温培养箱等;抗氧化酶活性和MDA检测试剂盒购自上海亚培生物科技有限公司;各试剂除甘油(中国国药上海化学试剂有限公司)、OEP(NOVA Chemical sales公司)外,葡萄糖、海藻糖、Tris及TES等均购自SIGMA公司.
1.2 精液冷冻与解冻
采用人工手握采精法,精液样品来自10头成年巴马香猪,采精频率为每周一次.所采集的精液用4层灭菌纱布过滤,去除其中的胶质,将精液转移至贮精瓶中.取富含精子部分的精液样品,只有活力大于70%的精液可用于试验.
取5mL精液于15mL离心管内,加入10mL精液低温保存液(BTS溶液),用12层纱布包裹并置于15 ℃恒温培养箱中预平衡3h;于15 ℃,500r/min离心5min,弃上清液,加入等温冷冻Ⅰ液(每100mL冷冻Ⅰ液由80mL基础液和20mL卵黄组成;精液冷冻基础液为每100mL三蒸水溶解Tris 2.42g,柠檬酸1.48g,果糖1.10g,5万IU的青霉素以及25mg庆大霉素),调整精子密度为1.5×109spz/mL,轻轻摇匀后,将12层纱布包裹的离心管置于5 ℃冰箱中降温平衡2h,用等温Ⅱ液(每100mL冷冻Ⅱ液由96mL冷冻Ⅰ液和4mL甘油组成)按照2∶1的体积比稀释,调整精子密度为1.0×109spz/mL,5 ℃的冰箱继续平衡1h;在5 ℃冰箱内,用专用注射器将精液迅速吸入等温的0.25mL细管,PVP粉末快速封管.
将细管和已预冷至-110℃的超低温温度计同时置于距离液氮面3cm处的冷冻架上,盖上盖子,于液氮上方熏蒸20min左右,待温度将至-120℃始冻温度时将细管迅速放入液氮中冷冻保存.解冻时将冻精细管(0.25mL)在37 ℃水浴中解冻45s,用2.25mL等温解冻液(95% BTS+5%冷冻Ⅰ液,v/v)稀释,37 ℃水浴孵育10min.
1.3 精子质量检测以及抗氧化酶(SOD,GPX)和丙二醛(MDA)的测定
本实验分别检测4种状态的精子质量和ROS水平.取各阶段精液15μL,使用计算机辅助精子分析仪(CASA)分析精子的运动参数,包括精子活率(TMS)、精子直线运动速率(VSL)、精子曲线运动速率(VCL);精子头部侧向移动振幅(ALH),实验重复5次,计算平均值;采用低渗肿胀(HOST)实验进行检测精子质膜完整性(PMI),将解冻后的精液用等温低渗液(7.35g柠檬酸钠和13.51g果糖溶于1L蒸馏水中制成)调整密度至1.0×106个,37℃培养30min,取10μL精液在血小板计数板上,400×倒置显微镜下观察不同部位的5个视野,计算尾部弯曲精子的百分率,质膜完整的精子因低渗导致膨胀表现为尾部弯曲;采用考马斯亮蓝染色法检测精子顶体完整性(AI),精子涂片用0.05%考马斯亮蓝(G250)染色,于1000×普通光学显微镜观察,顶体完整的精子顶体部分蓝染,顶体不完整的精子顶体不染色,每片计数200个以上的精子;使用SOD,GPX和MDA检测试剂盒,按照试剂盒说明书测定1×108个精子中相关物质活性及含量.
1.4 数据统计分析
通过SPSS 18.0统计软件,方差分析采用Duncan法进行多重比较;相关分析采用双尾法进行显着性检验.
2 结果与分析
2.1 巴马香猪精液冷冻降温过程中精子运动参数的变化
使用CASA系统分析巴马香猪精液冷冻降温过程中精子TMS,VSL,VCL和ALH等指标,以分析其运动参数的变化(表1).15 ℃精子各运动参数和鲜精均无显着差异(p>0.05);5℃精子的TMS和VSL均显着低于鲜精(p<0.05),ALH与15 ℃精子无显着差异(p>0.05);冷冻-解冻后精子的TMS和VSL显着低于其他各组(p<0.05),VCL显着高于其他各组(p<0.05),ALH与其他各组无显着差异(p>0.05).实验提示精子在降温至低于15 ℃后直至冷冻时的运动能力逐步降低,冷冻-解冻后的精子运动能力最低(p<0.05).【表1】
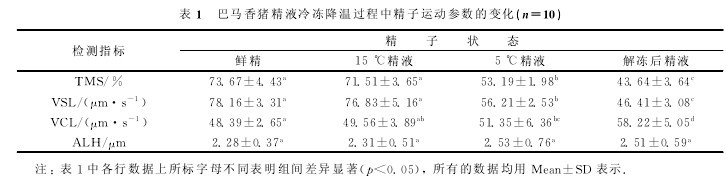
2.2 巴马香猪精液冷冻降温过程中精子细胞膜完整性的变化
通过HOST测试和考马斯亮蓝染色实验分别检测各降温阶段巴马香猪精子的PMI和AI,结果如图1所示.15℃精子的PMI和AI与鲜精无显着差异(p>0.05);5℃精子的PMI和AI均显着低于鲜精(p<0.05);解冻后精子的PMI和AI进一步降低,均显着低于其他各组(p<0.05).实验提示随着温度的降低,尤其是当精液温度低于15℃后,精子的质膜和顶体开始出现损伤,冷冻过程对精子的细胞膜损伤最为严重.
2.3 巴马香猪精液冷冻降温过程中精子抗氧化酶活性的变化
分别检测精液冷冻降温各阶段巴马香猪精子内SOD,GPX的活性,结果如图2所示.【图1-2】
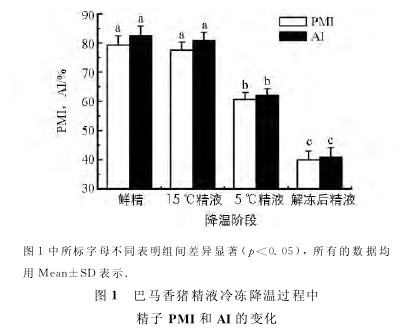
图3中所标字母不同表明组间差异显着(p<0.05),所有的数据均用Mean±SD表示.图3巴马香猪精液冷冻降温过程中精子活性氧水平的变化15 ℃精子SOD和GPX的活性与鲜精无显着差异(p>0.05);5℃精 子 的SOD活 性 低 于鲜精(p<0.05),GPX活性与 鲜精无显着差 异(p>0.05);解冻后精子的SOD和GPX活性均低于其他各 组 (p<0.05).
实 验 提 示 在 精 液 冷冻降温过程中,当精液温度低于15 ℃后,GPX活性受 低 温 影 响 较 小,SOD对 温 度 更 为 敏 感,而在冷冻阶段该两种抗氧化酶活性则均受到较大影响.
2.4 巴马香猪精液冷冻降温过程中精子活性氧水平的变化
通过MDA检测试剂盒检测精液冷冻降温各阶段巴马香猪精子内MDA水平,结果见图3.【图3】
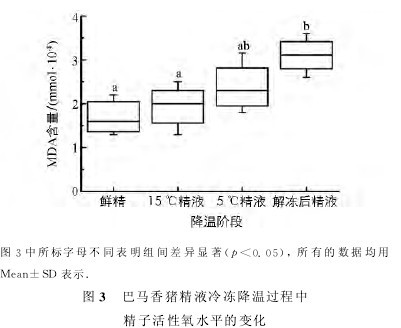
15℃精子和5℃精子的MDA含量与鲜精无显着差异(p>0.05),解冻后精子的MDA含量高于其他各组(p<0.05);各状态精子的MDA水平呈逐渐上升的趋势.实验提示当温度低于15℃后,随着温度的降低,精子内ROS水平逐渐上升,尤其是精液冷冻阶段,精子内的ROS水平达到最高.
2.5 巴马香猪精子抗氧化酶活性与细胞膜完整性的相关性分析
对巴马香猪各阶段精子中SOD,Gpx活性与PMI,AI的水平进行相关性分析,结果如表2所示.【表2】
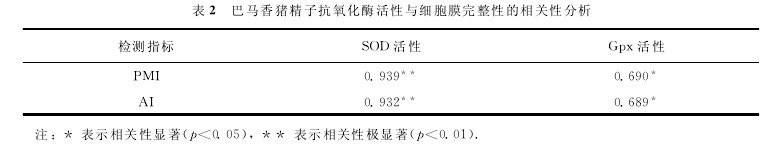
SOD活性与精子PMI和AI之间均具有极显着的相关性(p<0.01),Gpx活性与精子PMI和AI均具有显着的相关性(p<0.05).实验提示在巴马香猪精液冷冻的降温阶段,随着精子内抗氧化酶活性的降低,精子细胞膜受损程度也逐渐加大.
3 讨论
线粒体呼吸链中的一系列氧化还原酶是机体自由基产生的主要场所之一,而精子富含线粒体,ROS的产生也因此相当活跃.精子膜上富含不饱和脂肪酸,ROS能引发精子膜脂质的过氧化及其链式反应,从而导致精子膜受损.因此,精液冷冻过程中ROS的增高,是引起精子膜脂质过氧化,从而造成精子损伤的一个重要原因.机体存在抗氧化防御系统负责清除多余的ROS,通常情况下机体内ROS的产生和清除处于一种动态平衡.然而,当ROS过量产生,超过抗氧化防御系统的清除能力时,则会引起精子的应激损伤.
精液及精子中的酶促防御系统对清除自由基、保护细胞及机体起重要作用,该系统中酶活性的降低将会直接影响精子的品质.
本实验发现,巴马香猪精液降温过程中,15 ℃预平衡时,精子内抗氧化酶活性与MDA水平均未发生变化,精子没有出现明显的损伤(p>0.05);5℃平衡阶段,SOD和GPX水平显着下降,精子的运动能力和细胞膜完整性也显着降低(p<0.05),虽然该状态精子MDA水平与鲜精没有显着差异,但由于抗氧化酶活性的降低,可能导致了MDA以外的ROS成分的增加,从而引发精子细胞膜的脂质过氧化反应,导致精子质膜和顶体受损,并最终表现为精子活力下降.
与其他状态相比,冷冻-解冻后精子的运动能力和细胞膜完整性均显着降低(p<0.05),SOD和GPX水平最低,而MDA水平最高,可能是由于在冷冻过程中抗氧化酶活性过低,不能清除多余的ROS,从而导致了精子质膜和顶体受损,并引起精子品质的降低.
巴马香猪精液冷冻过程中抗氧化酶活性与精子细胞膜完整性密切相关,其相关性分析结果也证实了这一观点.当精子降温至15 ℃以下时,SOD和GPX活性开始降低,精子细胞清除ROS的能力逐渐下降,精子细胞膜脂质过氧化反应增强,PMI和AI也随之降低,细胞内容物外渗,精子受损;精子冷冻过程中,SOD和GPX活性降至最低,细胞内MDA水平显着增高,细胞膜受损程度加重,精子品质进一步降低.因此,通过研究抗氧化酶活性与精子细胞膜完整性之间的关系,本实验发现精液冷冻过程中除添加非酶促抗氧化剂辅助精子细胞抵御ROS的损害外,保护抗氧化酶活性也可能有助于提高巴马香猪冷冻精液品质.





