
疫苗在传染病防控领域发挥着至关重要的作用,极大程度降低了传染病的发病率和死亡率。可通过接种疫苗来抵御病原体入侵以预防疾病,如天花的根除、小儿麻痹以及甲乙型肝炎的控制等。但是随着疫苗种类的不断增多,不同疫苗之间存在接种禁忌,多个疫苗接种的程序繁琐等问题也随之引发。不仅造成资源浪费,而且增加感染机率,同时大量疫苗的储备及运输也提高了成本,由此给疫苗的使用和疾病的预防带来诸多不便。为解决上述问题,构建多联多价疫苗具有重要意义。目前,我国的麻疹-腮腺炎二联苗,麻疹-风疹二联疫苗以及麻疹-腮腺炎-风疹三联疫苗已成功研制[1-2]。对于烈性传染病,加强预防控制方案尤为重要。黄热病、汉坦病、克里米亚刚果出血热和炭疽等均是由病毒或细菌引起的以发热、出血、高病死率为特征的烈性传染病,对社会及人类健康具有极大的威胁。
痘苗病毒作为疫苗最早曾用于天花的防控,应用历史悠久。且痘苗病毒因其基因组容量大,含有大量的非必需基因,可载入大片段的外源基因等特点一直被广泛应用于构建基因工程病毒载体疫苗,如艾滋病[3]、流感[4]等。痘苗病毒天坛株(vacciniavirus Tian Tanstrain,VTT)是中国特有的痘苗病毒毒株,毒力相对较弱,具有高效表达蛋白、无致癌性、宿主细胞范围广等特点,适于多联多价疫苗的研究[5]。本试验以VTT为载体,构建含有4种外源基因的重组痘苗病毒,为多联多价疫苗的研制奠定了基础。
1、材料与方法
1.1细胞、病毒与质粒
BHK-21细胞,VTT由本实验室保存;质粒pSK-TC-EGFP(由上海捷瑞公司合成);pVaxⅠ-Cre(由本实验室构建并保存)[6]。
1.2主要试剂
胎牛血清购自HyClone公司,脂质体Lipofectin2000、OPTI-MEM购自Invitrogen公司,限制性内切酶购自NEB公司。
ExTaqDNA聚合酶、DL2000DNA Marker等均购自大连宝生物工程公司;细胞基因组提取试剂盒购自AXY-GEN公司;总RNA提取试剂盒购自上海生工生物工程公司。
1.3重组质粒的设计与合成
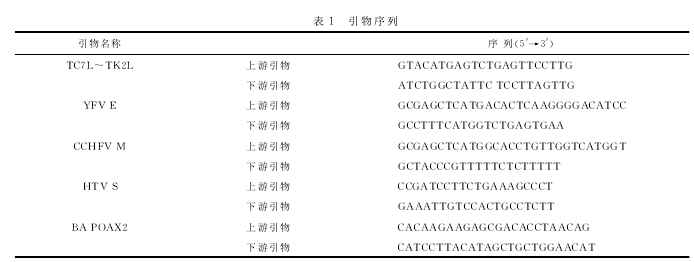
对痘苗病毒天坛株的基因组进行分析,设计TC7L~TK2L两侧重组臂,TC7L为天坛株痘苗病毒全序列从第14757~15248位,共492个碱基,TK2L为天坛株痘苗病毒全序列从第25469~26479位,共1011个碱基;结合痘苗病毒早晚期强复合启动子pE/L和特有的终止信号T5nT,以及外源筛选标记绿色荧光蛋白EGFP基因,构建包含4种外源基因的重组质粒pSK-TC-EGFP。每种外源基因两侧添加有酶切位点,均具有独立的表达框。该穿梭载体质粒由上海捷瑞公司合成。
1.4重组痘苗病毒rVTT-C-4+的构建及筛选
根据脂质体转染使用说明,将重组质粒与VTT共转染BHK-21细胞,步骤如下:用含2%血清的DMEM培养液培养BHK-21细胞,取1×105个细胞接入6孔板中,于37℃、5%CO2条件下培养24h,以0.1个感染复数(multiplicity of infection,MOI)的野生型VTT感染BHK-21细胞,感作2h后用重组质粒共转染;培养72h后反复冻融细胞3次以收获病毒,将收获的病毒再次接种于BHK-21细胞,并于1%甲基纤维素环境中培养72h,于荧光显微镜下挑取绿色荧光蚀斑;重复上述筛选过程,直到获得纯化的含EGFP基因的重组痘苗病毒rVTT-C-4+EGFP+。按上述方法将质粒pVaxⅠ-Cre与获得的rVTT-C-4+EGFP+共转染BHK-21细胞,并于1%甲基纤维素环境中培养72h,在荧光显微镜下挑取无绿色荧光的蚀斑;重复上述筛选过程以获得重组痘苗病毒rVTT-C-4+。
1.5重组痘苗病毒rVTT-C-4+的PCR鉴定
提取重组痘苗病毒rVTT-C-4+的基因组为模板,用表1所示5对引物进行PCR。其中CCHF的M基因的反应条件为95℃5min;预热95℃30s,退火54℃30s,延伸72℃30s,30个循环;72℃延伸10min;其余4对引物的退火温度均为58℃,延伸时间1min10s。分别以VTT基因组和重组质粒作为阳性对照,引物序列见表1。
1.6重组痘苗病毒rVTT-C-4+的形态学观察
利用负染法对重组痘苗病毒rVTT-C-4+进行形态学观察。用rVTT-C-4+感染BHK-21细胞,待细胞完全病变后,收集病变细胞,反复冻融3次,3000r/min离心5min,取上清用2%磷酸钨溶液(pH6.8)处理30s,置于电镜下观察。
1.7重组痘苗病毒rVTT-C-4+遗传稳定性的检测分别以重组痘苗病毒第5、10、15、20代重组痘苗病毒的基因组为模板,用如表1所示5对引物进行PCR。其中CCHF的M基因的反应条件为95℃5min;预热95℃30s,退火54℃30s,延伸72℃30s,30个循环;72℃延伸10min;其余4对引物的退火温度均为58℃,延伸时间1min10s。分别以VTT基因组和重组质粒作为阳性对照。
2、结果
2.1重组质粒的设计及合成
选取了XhoⅠ、KpnⅠ、SacⅠ、NotⅠ、XbaⅠ、SalⅠ、SmaⅠ和SpeⅠ8个酶切位点,分别置于4种外源基因的两侧。重组质粒pSK-TC-EGFP的线性构建图如图1所示,包括两侧重组臂、启动子、终止信号、酶切位点、外源筛选标记基因等。
EGFP编码绿色荧光蛋白,用于重组病毒的筛选,且EGFP表达盒外侧携带Loxp序列,便于EGFP基因的敲除。
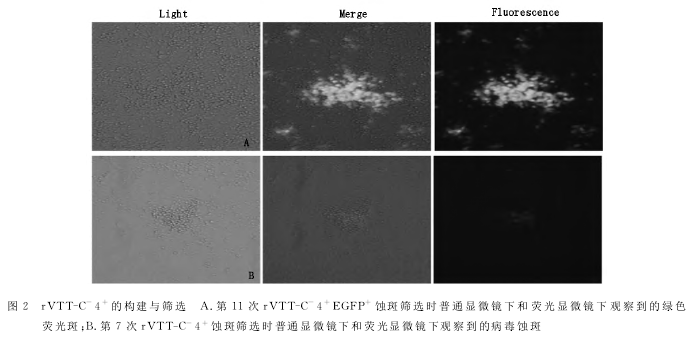
2.2重组痘苗病毒rVTT-C-4+的构建与筛选
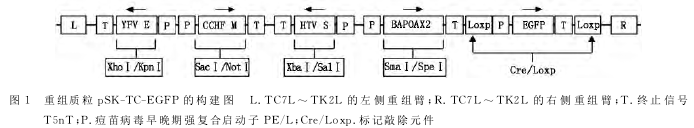
重组质粒与VTT共转染BHK-21细胞后,经17次含绿色荧光的蚀斑筛选得到纯化的重组痘苗病毒rVTT-C-4+EGFP+,获得的rVTT-C-4+EGFP+利用同源重组与质粒pVaxⅠ-Cre共转以敲除EG-FP基因,经10次无绿色荧光的蚀斑筛选,得到纯化的重组痘苗病毒rVTT-C-4+。图2A所示为rVTT-C-4+EGFP+第11次蚀斑筛选时普通显微镜下和荧光显微镜下观察到的绿色荧光斑,图2B所示为rVTT-C-4+第7次蚀斑筛选时普通显微镜下和荧光显微镜下观察到的病毒蚀斑。
2.3重组痘苗病毒rVTT-C-4+的PCR鉴定
结果显示,图3A以VTT基因组为模板扩增出1条300bp的条带,以重组痘苗病毒为模板的泳道内无条带,说明重组痘苗病毒TC7L~TK2L基因已完全敲除;图3B重组痘苗病毒为模板扩增出1条250bp的条带;图3C重组痘苗病毒为模板扩增出1条250bp的条带;图3D重组痘苗病毒为模板扩增出1条500~750bp的条带;图3E重组痘苗病毒为模板扩增出1条500bp的条带;与预期结果相符,说明重组痘苗病毒构建成功。
2.4重组痘苗病毒rVTT-C-4+的形态学观察
利用负染法观察病毒形态特征,电镜下痘苗病毒呈砖型或卵形,约为200nm,在成熟病毒表面有规则的条形或颗粒状的突起。图4A为野生型痘苗病毒VTT的电镜图,图4B所示为重组痘苗病毒rVTT-C-4+的电镜图,通过图片显示重组痘苗病毒与亲本相比形态无显著差异,说明外源基因载入对于痘苗病毒的形态形成无显著影响。
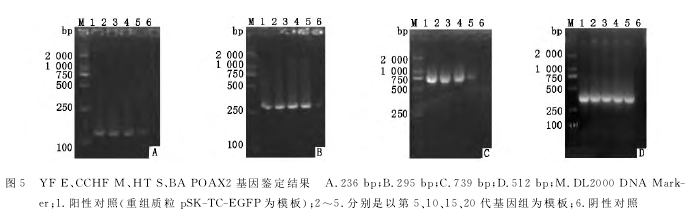
2.5重组痘苗病毒rVTT-C-4+遗传稳定性的检测
利用PCR对重组痘苗病毒的遗传稳定性进行分析,结果如图5所示,在第5、10、15、20代的重组痘苗病毒均扩增出4段外源基因,说明构建的重组痘苗病毒具有良好的遗传稳定性。
3、讨论
多联多价疫苗为同时防治多种疾病提供了可能,减少了接种次数,简化了免疫程序,提高了免疫效率,同时降低了多种疫苗在研发、生产和运输过程中的成本,以免造成不必要的资源浪费和疫苗价格的提高。结合多联多价疫苗所具备的优越性,本试验利用基因工程技术构建重组痘苗病毒多联多价侯选疫苗。以痘苗病毒为载体的活疫苗有如高效、廉价、易于构建多联疫苗等多个优点。但痘苗病毒作为疫苗载体在应用上仍然存在一些不足之处:其一是在接种后会出现较严重的局部反应,甚至可能引起严重并发症;其二是机体会产生针对痘苗病毒的中和抗体,会影响再次接种相应载体疫苗的免疫效果。近年来,利用基因工程对痘苗病毒不断改造,为解决以上问题带来了希望。如经由非敏感宿主体内多次传代而产生的改良型安卡拉毒株(MVA);此外,在痘苗病毒哥本哈根株基因组中敲除非必需基因区域的TK、HA、ATI基因等18个基因而构建的弱毒株NYVAC[7]。虽然这些非复制型载体具有优越的安全性,但在免疫过程中需要多次接种高滴度的病毒载体才可诱导较强的免疫应答。本试验将痘苗病毒天坛株基因组中的TC7L~TK2L基因敲除,同时插入4种外源基因,构建多联重组痘苗病毒疫苗。根据缺失痘苗病毒相关基因的研究[7]以及Kan等[8]前期研究结果,该基因的缺失可导致痘苗病毒的毒力减弱。所以本试验本着减弱毒力而不影响病毒复制和基因表达的前提下,构建多联重组痘苗病毒。
黄热病病毒(YEV)E蛋白是重要的跨膜蛋白,是诱导中和抗体的主要抗原[9-12]。克里米亚-刚果出血热病毒(CCHF)的基因组由大(L)、中(M)、小(S)3个节段组成,其中M基因编码糖蛋白(GP)[13],成熟的糖蛋白可以识别易感宿主细胞上的受体并能诱导中和抗体产生[14]。汉坦病毒(HT)基因组中的S基因编码其核蛋白(NP),NP在该病毒结构蛋白中免疫原性最强,并含有B细胞和T细胞表位,可刺激机体产生体液免疫和细胞免疫[15-17]。炭疽杆菌(BA)的POAX2基因为表达荚膜质粒的基因,而荚膜抗体可以诱导调理机体对炭疽芽孢杆菌的吞噬作用,抗荚膜的抗体可诱导机体清除炭疽芽孢杆菌的繁殖体,从而保护机体抵抗炭疽感染[18]。以上4段基因均为现在应用比较成熟的基因,联合以上4种外源基因,将其插入痘苗病毒基因组中,构建四联重组痘苗病毒。
对于一个安全有效的活载体多联疫苗具有遗传稳定性是必要的,为了克服多种外源基因在表达时可能会发生相互竞争和干扰,每种外源基因都设计了独立的表达盒,且具有各自的启动子和转录终止信号。另外,我们采用强效复合型启动子(pE/L)来提高外源基因的表达效率,虽然痘苗病毒中最强的启动子为痘苗病毒晚期启动子,但一般情况下它所表达的抗原只产生中和抗体[9],所以正确选择不同时期启动子表达外源基因是重要的。痘苗病毒早晚期pE/L强效复合型启动子表达外源基因可诱导良好的体液免疫和细胞免疫。在重组毒株的筛选方面,本试验选择以表达绿色荧光蛋白(EGFP)作为筛选标记,因其荧光稳定性好、片段小容易构建、方便观察,同时与传统方法相比解决了筛选周期长,筛选程序复杂等问题,所以本试验应用EGFP对重组病毒进行标记。在获得重组毒之后,为了后续研究,在EGFP两端设置Loxp序列,以便通过Cre/Loxp系统将标记基因删除。本试验将4种烈性传染病的病原基因载入痘苗病毒基因组的非必需区域,同时利用基因重组技术缺失痘苗病毒的相关毒力基因TC7L~TK2L,图4表明4种外源片段的载入对痘苗病毒的形态形成无显著的影响。图3表明成功构建含4种外源基因的重组痘苗病毒,并在转录水平上检测了病毒连续传10代的遗传稳定性,说明4种外源基因均可在痘苗病毒中进行稳定的表达,可应用于后续的研究。本研究所针对的4种外源基因均为危害人类健康及公共安全的烈性传染病,由于相应抗体的限制,本试验未针对表达的蛋白进行进一步检测,但是病毒的构建是多联疫苗研制的基础。
因此,本试验旨在构建四联重组痘苗病毒,为多联多价疫苗在预防和控制疾病的发生和临床应用方面奠定了基础。
参考文献:
[1]储艳,陆玉忠,陶红,等.国产麻腮风联合减毒活疫苗的安全性及免疫原性[J].中国生物制品学杂志,2011(6):707-710.
[2]梁祁,周伟忠,李靖欣,等.麻疹风疹联合减毒活疫苗的安全性及免疫原性观察[J].中国生物制品学杂志,2011,24(12):1490-1493.





