第一章 绪 论
1.1 黄瓜白粉病及其抗性基因定位研究概况
黄瓜(Cucumis sativu. L.)是我国主要的蔬菜作物,是面积最大的保护地栽培蔬菜之一。黄瓜白粉病是一种为害严重的世界性病害(冯东昕 等,1996;Block et al,2005),在生产上,白粉病与霜霉病、枯萎病并称为黄瓜三大病害。白粉病在我国各地均有发生,且常发生在结瓜盛期,导致叶片光合能力下降或者丧失,严重影响果实的品质和产量。
为了应对白粉病的发生,研究者对黄瓜白粉病的发病规律和抗性基因的定位方面进行了大量的研究,取得了一系列进展,特别是随着国际黄瓜基因组计划的开展(张圣平 等,2010),黄瓜全基因组测序的完成,开发了大量的新型分子标记(SSR,InDel 等),大大促进了白粉病抗性基因的研究工作,但即使如此仍然没有得到一致的结论(刘苗苗 等,2009)。这主要是由以下三个因素导致的:首先,研究材料的差异,不同的研究材料带有的抗性基因不同,抗性机制也可能不同,抗性的程度也有差异,导致不同的学者研究结果分歧很大;其次,白粉病的发生和发展受周围环境(如温度、湿度等)影响较大,难以保证抗病性鉴定的发病条件完全一致;第三,目前对白粉病抗性的鉴定主要是通过研究者肉眼观察发病情况,对其抗性进行评价,而不同的研究者的评价标准具有很大的主观因素干扰。
1.1.1 黄瓜白粉菌简介
白粉病的病原菌是真菌白粉菌科,菌丝全部或大部分暴露在寄主植物的茎、叶和果实等表面,产生大量的菌丝体和分生孢子等白色粉状物,故而得名,白粉病在全世界范围内广泛分布。由于白粉病的巨大危害,世界瓜类生产国都将白粉病抗性育种列为主要育种目标之一(刘秀波 等,2005)白粉菌寄主范围因病原菌种类不同而异,但是一般的来说,一种植物只发生一种白粉病,但也有例外。黄瓜白粉病原菌主要有两个属:单囊壳白粉菌 Podosphaera xanthi(i异名(Sphaerothecafuliginea))和二孢白粉菌 Golorinomyces cichoracearum(异名 Erysiphe cichoracea rum)(张素勤 等,2007;刘龙洲 等,2008),在中国主要是前者(Xu et al.,1999;王建设 等,2003;张艳菊 等,2010)。该病原菌寄主范围主要集中在黄瓜、西甜瓜、南瓜等葫芦科作物,并有多种生理小种(曲丽 等,2007)。白粉菌孢子在 15-30℃下均可萌发,以 20-25℃最适。对湿度要求更广,最低在相对湿度 25%下也可以萌发,但是在过高的湿度下容易吸水破裂,45~75%时发病快,超过 95%受到抑制。在做白粉病抗性的人工鉴定时,一定要注意保持白粉病发病的适合条件才能得到准确的结果。
1.1.2 黄瓜白粉病抗病性鉴定方法
对于作物的基因定位研究而言,表型鉴定是至关重要的一个环节,而抗病基因的定位研究就需要对作物的抗病性进行客观的、准确的、稳定的评价。影响黄瓜白粉病发生和发展的因素很多,不同的接种方法、环境条件的控制都会直接影响鉴定的结果。为了使不同的研究人员的结果具有可比性和比较好的重复性,有必要对抗病性鉴定的方法进行一些科学的规范。下面介绍一下本实验采用的抗病性鉴定方法(参考中华人民共和国农业行业标准 NY/T 1857.2-2010 第 2 部分:黄瓜白粉病鉴定技术规程)。
1.1.2.1 苗期孢子悬浮液喷雾接种鉴定技术
该技术是指在植株苗期时,从已发病叶片上刷取白粉菌孢子到清水中,加入 SDS 后高速搅拌10min,配置成孢子悬浮液,然后将悬浮液均匀的喷洒到待鉴定植株叶片上。白粉病苗期孢子悬浮液喷雾接种鉴定比其他的接种鉴定方法具有非常明显的优势:实验用地少、周期短、方便操作,大大减少了外界环境的影响的同时,一季度可以同时进行多次重复鉴定,加速鉴定进程和可靠性。多数研究者选用此方法(如张圣平 等 2011;刘龙洲 等 2008)。
黄瓜白粉病苗期抗性与成株期抗性的关系的相关报道不多,但是有多位学者对黄瓜霜霉病的苗期抗性与成株期抗性进行过研究报道,Jenkins(1946)对黄瓜霜霉病的抗性做了田间和室内的对比,认为苗期抗性与与成株期抗性表现一致。1994 年,我国学者许启新对 9 种材料进行了研究,发现其苗期接种和成株期自然发病情况的相关性达到极显着水平。黄瓜霜霉病和白粉病一样都是真菌性病害,且已有研究表明黄瓜的白粉病抗性与霜霉病抗性具有显着相关性(Van Vliet et al.,1977;丁国华 等,2004)1.1.2.2 田间成株期抗病性鉴定技术。田间成株期鉴定是指在病圃中正常栽培管理植株,让其自然发病,待其充分发病后进行病情的调查。这种方法的前提是种植的病圃需要在往年连续发病,可为鉴定提供充足的侵染源,也有多位学者采用这种鉴定方法(如张素勤 等,2005;程嘉琪 等,2011)。田间成株期充分发病后的调查方法不同于苗期接种鉴定,因为发病时底部的叶片或已衰老脱落,不能反应其真实抗性,所以一般调查从下往上第 3~8 片真叶的病级的平均数作为这一株的病级,然后计算病情指数。这样鉴定的结果更接近于自然发病,更符合生产上的环境和条件,能更好的说明其在实际生产中的抗性。
1.1.2.2 叶盘接种鉴定
所谓叶盘法就是用特定大小的圆形模具在黄瓜真叶上截取相同大小的圆盘,放在保湿的培养箱中,维持叶片正常的生理状态,然后进行悬浮液喷雾接种,让其在离体的情况下发病。发病后调查病斑面积,以其占圆盘面积的比例作为其发病情况的依据(Fukino et al., 2013)。这种方法的优点是节省空间,省时省力,容易控制环境因素的影响。缺点是截取叶片时会对叶片造成一定程度的损伤,破坏叶片原有的代谢方式,可能会改变很多酶的代谢活性或者表达量,而这些变化很有可能直接影响病原菌的入侵,进而影响鉴定结果;同时由于伤口的存在,叶片容易受到环境中细菌的感染,使叶片腐烂,影响白粉病斑的扩展和观察,也会影响抗病性鉴定结果。
1.1.3 黄瓜白粉病发病规律及防治方法
黄瓜白粉病在植株全部发育阶段均可发病,一般是中后期较重,主要侵染叶片,影响叶片的光合功能,甚至丧失光合能力,茎部和果实也可发病,但一般较轻。发病过程如下:初期产生白色斑点,环境条件适宜时向四周扩展至连成片,最后整个叶片布满白色霉层。病害一般是从下往上侵染,发病后期,叶片枯黄,坏死,加速植株衰老(沈丽平 等,2009)。
防治白粉病的措施有很多,如加强田间管理、对温室进行消毒处理等,但在生产上,防治白粉病主要依靠农药防治(周益林 等.2001),为了达到较好的防治效果,一般需要过量用药,增加了生产成本的同时也对环境造成巨大污染。所以培育抗白粉病新品种是防治白粉病最经济最有效的手段。为了培育抗病新品种,研究人员对黄瓜白粉病抗性的遗传规律、与抗性基因连锁的分子标记开发、抗病基因定位等方面进行了大量的研究。
1.1.4 抗性遗传规律研究
前人已有多篇关于黄瓜白粉病抗病遗传规律的报道,但是不同学者研究的结果差异很大,尚不统一,总结现有报道可归纳为以下 4 种观点:
(1)隐性多基因控制的数量性状:1948 年,P. G. Smith 利用抗性材料 Puerto Rico 37 与感病材料杂交,发现 F1趋向于感病,在后代 F2中的抗感比为 1:21,认为 Puerto Rico 37 中的白粉病抗性是由多个隐性基因控制。1956年,Barnes 等以抗病材料 PI197087 为研究材料,通过构建遗传群体进行遗传分析,认为该抗性材料中含有一或两个主效基因和一或两个微效基因功能控制。1995 年,吕淑珍等通过研究认为黄瓜中最少 3 个以上的基因其对白粉病的抗性,并且感病表现为显性性状,具有较高的遗传力。
Kooistra(1968)用三个亲本构建两个遗传群体,分别进行配合实验,得出黄瓜对白粉病的抗性在两份不同的抗病材料中分别受到不同基因的控制:Natsufushinari 中含有 2 个抗病基因而 PI2008151 中只含有 1 个隐性的抗病基因,这三个基因均不相同,在 F2后代中表现出超亲现象。张素勤(2005)用三个抗感杂交组合进行遗传研究,认为有 2 个主基因控制黄瓜白粉病抗性,并且遗传率均很高。
(2)隐性单基因控制:腾枝国光(1962)认为“青节成”中存在一个隐性的抗病基因。用这种抗病材料与感病材料杂交,F1均表现为感病,F2中抗感分离比为 1:3,依此得出其抗性有一个隐性的单基因控制。张桂华(2004)也通过构建遗传分离群体进行遗传研究,得到了与其相同的结论。刘龙洲(2008)用一对抗感亲本构建遗传群体,进行了苗期接种鉴定,结果后代分离群体(F2、BCR 和 BCS 群体)中分离比的结果完全符合孟德尔遗传规律(P≤0.05)。由此得出结论,在抗病亲本中由 1 个隐性基因控制其抗性,感病性状为不完全显性。
(3)由一对不完全隐性基因和 2 对上位修饰基因控制,Shanmugasundaram 等(1971,1972)用对白粉病具有完全抗性的黄瓜种质材料 P1212233、P123514 分别与感病亲本进行杂交,发现 F1均表现为感病,F2群体中表现为抗性性状分离,并且高抗、中抗、感病的单株数之比符合 1:3:12,根据孟德尔遗传分离定律,认为其抗性是由一个隐性主基因 s,一个显性基因 R 和一个显性抑制基因 I 控制。这三个基因各自特点如下:s 是主效基因,决定下胚轴抗性,同时也是叶片抗性所必需的;R 基因主要影响叶片的抗性,I 基因为抑制基因,可使植物表现出不完全抗性,但对 s 基因的表达不产生影响。随后,毛爱军等(2005)、简德明(2007)、王振国(2007)等分别以 WI 2757(高抗白粉病,美国陆地黄瓜自交系)与我国一份栽培品种进行杂交配置 F2为材料进行遗传分析,研究结果与 Shanmugasundaram 基本一致。
(4)由一对隐性基因和一对不完全显性基因控制,与温度有关Morishita 等(2003)发现黄瓜的白粉病抗性由一对隐性基因和一对不完全显性基因控制,并且抗性受到温度的调节。根据其抗性对不同温度条件的反应不同,将黄瓜材料分成三种类型:高温(26℃)和低温(20℃)下均表现为抗性的品种,如 PI197088-5(P)等,在高温和低温下均表现为感病的品种,如 Sharp(S),第三类就是只在高温下才表现出抗性的品种,如 Natsufushinar(iN)。
用这三种类型的黄瓜进行互相杂交、自交和回交,并分别进行抗病性鉴定,结果表明:有两个基因控制黄瓜的抗白粉病性状 aa 和 BB。只有 aaBB 这种基因型时,黄瓜才能在高温和低温下均表现出抗性,且为完全抗性;当为 A_bb 时,黄瓜在高温和低温下均表现出感病;当为 aaB_时,黄瓜会表现出高温下抗病而低温下感病。
可以看出,黄瓜的白粉病抗性机制复杂,黄瓜白粉病的抗性规律研究很多而杂,尚未得出统一结论,这严重影响下一步白粉病抗性基因的进一步精细定位和克隆。一般认为主要由以下几个因素造成:首先黄瓜白粉病病原菌具有不同生理小种,不同的研究所用的病原菌有差异;其次,不同研究者进行抗病性鉴定采用的鉴定方法不统一,如接种时机、环境条件控制、病情调查时间等,并且多为肉眼观测,易引入主观误差,不同的抗病材料所含有的抗病基因可能不同,抗病机制复杂。所以,不同学者的研究结果千差万别。
1.1.5 抗性分子标记与遗传定位的研究
在黄瓜的抗白粉病育种工作中,对材料的抗病性鉴定是影响育种进程最重要的因素,影响白粉病的田间鉴定的环境条件较多,费时费力,一些学者就试图通过寻找与抗病基因连锁的分子标记来进行分子辅助选择育种(MAB),从而提高育种效率。
张桂华(2004)利用 F2分离群体,结合 BSA 建池法构建抗感池,得到一个 AFLP 标记,并成功转化成 SCAR 标记,连锁距离为 5.56cM。随后,多位学者也陆续得到了多个与黄瓜白粉病抗性相关基因连锁的分子标记(张素勤 等,2005;简德明 等,2007;王振国 等,2007;张海英 等,2006)。这些分子标记一般都是 AFLP 或 SCAR 标记,稳定性较差且遗传距离相对较远。
张海英(2008)采用 SSR 分子标记技术,获得两个与黄瓜抗白粉病基因连锁的 SSR 标记,遗传距离最近达到了 5cM。聂京涛(2011)利用 BSA 建池法获得一个与白粉病抗病基因连锁的 SSR 分子标记 SSR15592,遗传距离为 7.62cM。随着分子生物学的发展,以及黄瓜抗病育种的需求,对白粉病抗性基因的定位研究也愈加深入,特别是 QTL 分析方面,取得了非常不错的成果:Sakata(2006)利用高抗材料 PI197088-1 和高感材料 Santou 作为亲本,构建了重组自交系(RILs)群体,共检测到 5 个与抗白粉病有关的 QTL,其中只有一个位点在高温(26℃)、低温(20℃)下对抗性均起作用,其他的仅在高温或者低温下起作用。这是第一次对不同温度条件下的抗病基因进行分别 QTL 定位。
刘龙洲(2008)利用黄瓜抗病自交系 S06 与感病自交系 S94 配置群体,构建了一个 F6:7重组自交系,分别与 2005 年秋和 2006 年春两个季度进行了苗期喷雾接种抗病性鉴定和 QTL 定位分析,认为白粉病抗性属于数量遗传,并且鉴定出至少四个 QTL 位点调控其抗性。这四个 QTL 位点分别命名为 pm1.1, pm2.1, pm4.1 和 pm6.1,其中 pm1.1、pm2.1、pm4.1 在两次实验中均被检测到。两季度检测到的 QTL 的表型解释率合计分别达到了 52.0%和 42.0%。检测到的位点较多,表型解释率也较高,但是,这些位点却没能与染色体对应。
张圣平(2011)利用从国内栽培品种中筛选出来的高抗和感病自交系作为亲本,构建遗传分离群体,进行遗传和 QTL 定位分析,认为其 F2后代群体中抗病性呈连续分布,符合数量性状遗传特征,最终检测到 4 个 QTL 位点(pm5.1、pm5.2、pm5.3、pm6.1),分别位于黄瓜 5 号和 6 号染色体上,其中,除了 pm6.1 之外,其余三个位点均被重复检测到,特别的 pm5.2 的表型解释率最高,在两次实验中分别达到了 36.1%和 41.6%,对应的 LOD 值分别达到了 15.06 和 17.88,检测到多个有价值的 QTL 位点,并且与染色体物理位置对应,具很高的应用价值。Fukino(2013)同样利用高代重组自交系进行了白粉病的 QTL 定位,用不同的鉴定方法,在不同的季度,不同的温度下进行了抗病性鉴定,检测到多个 QTL 位点,其中被重复检测到的表型解释率较高的 QTL 位点有:pm1.2、pm5.2、pm5.3、pm6.1,其中 pm1.2 和 pm6.1 在四次实验中均被检测到,pm5.3 被检测到三次。
He 等(2013)利用 F2群体,在不同季度下对子叶、下胚轴、真叶分别进行抗病性调查,并进行了 QTL 分析,并对前人的研究成果进行了详细的整合。发现除了黄瓜 2 号染色体上之外,其余的六条染色体均被检测到 QTL 存在。其中检测到次数最多的位点位于黄瓜 5 号染色体(Chr5)上。并且发现黄瓜下胚轴的白粉病抗性是由单基因控制,并且定位于 5 号染色体上。
综上所述,黄瓜白粉病抗性遗传机制复杂,检测到的相关基因较多,不同抗病材料所具有的抗病基因不同,并且分布在不同的染色体上。目前研究的结果主要集中在黄瓜 5 号染色体上,而其他染色体上的抗病基因研究较少,完成全基因组范围的白粉病抗性基因的挖掘将有利于综合利用黄瓜种质中的抗性资源。
1.2 黄瓜白粉病抗病基因挖掘的研究
1.2.1 植物病原互作模式的研究进展
在自然界中,充满了各种病原菌,包括真菌、细菌和病毒等,这些病原菌与植物的互相作用就会导致植物疾病的发生,如白粉病就是由于真菌(白粉菌)入侵植物体造成的。所以植物发病从根本上讲是病原菌与植物体之间的互相作用的外在表现。研究发现植物与病原互相作用的模式主要有两种:基因对基因假说和防卫假说。
基因-基因假说:1971 年,Flor 提出病原与抗病基因分别编码激发子和激发子受体,若激发子与其相应的受体识别即可激发植物的抗病反应,表现为抗病,否则表现为感病。这种抗性一般表现为显性。防卫假说:该假说认为,病原物释放一种效应因子,这些效应因子可以作用并修饰植物的某些目标蛋白,植物检测到这种修饰之后可以触发植物对病原的抗病性。防卫假说的提出改变了之前对抗病基因功能的认知,抗病蛋白不仅仅是被动的等待与病原蛋白结合,而是主动监视自身的生理变化,迅速启动防卫反应(Axtell et al., 2003;Martin et al.2003;王明 2012)1.2.2 植物抗病基因的类型。
植物中克隆的第一个抗病基因是于 1992 年在玉米中克隆出来的,至今已有 10 多种植物中的70 多个抗病基因被克隆出来(Liu et al.,2007),通过对这些已克隆的基因的研究发现,大多数抗病基因中存在保守的序列:如富含亮氨酸的重复序列(LRR),核苷酸结合位点结构域(NBS),CC(coiled-coiled)结构域等,根据其主要结构域把抗病基因分成了以下几个大类:
1、NBS-LRR 类抗病基因,这类抗病基因含有 NBS 和 LRR 的结构域。(Hammond-Kosack andParker, 2003)2、胞外 LRR 受体类,其编码蛋白包含胞外 LRR 结构域和单个跨膜区,胞内不编码激酶。甜菜中的 HS1 基因(Cai et al., 1997)3、拟南芥中发现的抗白粉病基因 RPW8,其编码产物为一个携带 CC 结构域的膜蛋白(Xiaoet al.,2001)4、大麦抗禾柄锈菌基因 Rpg1,其编码产物为一个带有两个激酶结构域的受体激酶类似蛋白(Brueggeman et al., 2002)5、编码胞内丝氨酸/苏氨酸蛋白激酶,这类抗病蛋白包含丁香假单胞杆菌基因 Pto 和 PBS1,这两个基因分别来源于番茄和拟南芥,属于不同的亚家族。6、编码胞外 LRR 类似受体激酶,编码的蛋白一般包含胞外的 LRR 结构、单个的跨膜区和胞内的丝氨酸/苏氨酸蛋白激酶(Song et al.,1995; Gomez-Gomez et al., 2000; Sun et al.,2004)7、Mlo 基因:这是从大麦中克隆的一类抗白粉病的隐性基因。其一般包含 7 个跨膜区,其显性基因的功能可能是抑制细胞程序性死亡,突变后几乎对所有白粉病小种具有抗性(Buschgesetal.,1997)。
综合以上几种类型的抗病基因,最重要的是 NBS 类抗病基因。NBS 结构中包含三个关键基序:P-loop、Kinase2 和 Kinase3a。具有结合 ATP 或 GTP 及水解酶的活性。NBS 结构是基因内互作的关键部位,同时也参与不同基因同源结构互作(Rathjen and Moffett, 2003)。NBS 结构突变可显着影响抗病基因的功能,例如 P2 基因的 NBS 结构内保守序列突变后,P2 基因基本丧失抗病功能(Dodds et al., 2001)。其次是 LRR 结构域,LRR 结构不尽是抗病蛋白与病原基因产物作用的部位,也决定抗原基因抗性专一性,在信号传导中也有重要的作用,LRR 结构的突变也会显着影响基因的抗病功能(Dinesh-Kumar et al.,2000;Bryan et al., 2000;Anderson et al.,1997)。有学者推测LRR 区是抗病基因产物和病原基因产物直接或间接作用的部位(Dangl and Jones,2001 )。He etal(2000)通过实验证明 LRR 决定抗病基因特异识别的激发子,而胞内激酶决定信号途径。
1.2.3 黄瓜抗白粉病基因挖掘的研究
虽然学者们对黄瓜白粉病抗性基因进行了大量的 QTL 定位实验,但是由于定位区域依然很大,缺少有效的手段进行进一步的精细定位,随着生物信息学的发展,有的学者开始利用基因的同源性,预测黄瓜中的抗白粉病基因。Yang 等(2013)在黄瓜中预测了 67 个 NB-LRR 类型的抗病相关的同源基因。这些基因分布在黄瓜 7 条染色体上,其中可能有部分与白粉病抗病性有关Schouten(2014)利用拟南芥中已经发表的白粉病抗性相关基因和黄瓜基因组测序数据,在全基因组范围内搜寻白粉病抗性基因,最终鉴定出 13 个 MLO-like 基因,10 个 AtPMR4 同源基因,一个 PMR5 同源基因,13 个 PMR6 同源基因。这些基因也是今后挖掘和验证白粉病抗性候选基因的重点。
1.3 黄瓜白粉病抗性基因全基因组关联分析(GWAS)
全基因组关联分析(Genome-WideAssociation Study)是一项最早应用于人类疾病研究的技术,Science 杂志于 2005 报道了第一项具有年龄相关性的黄斑变性的 GWAS 研究(Klein,2005),随后,陆续发表了一系列应用 GWAS 研究人类疾病的研究成果。随着测序技术的发展,测序技术的成熟和测序成本的降低,一些重要模式生物和作物已经完成全基因组测序和重测序工作,所以,一些针对重要的动植物进行的 GWAS 研究也陆续进行。
1.3.1 关联分析的优势
植物中第一个应用关联分析研究的性状是玉米中开花时间的基因 dwarf8(d8),并随后在拟南芥和其他作物中得到广泛应用。关联分析具有如下优势:(1)不需要构建遗传分离群体,缩短了研究时间并节约了成本;(2)连锁作图一般只有两个亲本,只能定位两个等位基因,而关联分析可以实现对一个基因座上多个等位基因的同时研究;(3)由于关联分析使用的是自然群体,所依据的重组是长期进化积累下来的,相对于遗传群体短时间内的有限次的重组,定位结果的分辨率更高,甚至可以直接找到基因;(4)关联分析使用的标记距离都是以 bp 为单位,直接与物理位置对应,可以更加容易的将定位结果与基因进行整合。(Liu et al.,2011;王明,2012)关联分析以长期重组交换保留下的位点之间的连锁不平衡(linkage disequilibrium, LD)为基础,获得群体表型数据和基因型数据后,用统计学方法检测位点多态性与性状遗传变异之间的关联,也可称之为连锁不平衡作图(Flint-Garcia et al., 2003),LD 的衰减程度决定了关联分析的分辨率。LD 衰减较快,关联的分辨率高,定位基因的准确性也高,相反,LD 衰减慢,精度降低,定位区间会相应的加大。
1.3.2 连锁不平衡与遗传连锁的关系:
经常会有人将连锁不平衡与遗传连锁两个概念混淆,实际上这是两个完全不相同的概念,遗传连锁是通过计算两个位点之间的重组率的大小来衡量,考虑的是两位点之间在染色体上的距离的问题或者说是分布的状态;而连锁不平衡分析考虑的是不同的位点之间的相关性,只要两个位点相对应的等位变异同时出现的概率大于群体中随机组合的概率,就称之为连锁不平衡。由于如果两个位点在染色体上位置较近,会导致其在群体中更容易同时出现,导致连锁不平衡,这也是导致人很混淆这两个概念的主要原因。连锁不平衡主要来自于两个因素:自然选择和基因漂变,但是紧密的连锁也会增加连锁不平衡性。连锁不平衡的度量本质上是实际观测到的单倍型频率与随机分离时的期望频率之间的差异。
1.3.3 植物中影响 LD 的主要因素
作物的交配系统是影响 LD 的主要因素。异交作物的 LD 衰减远小于自交作物。例如玉米栽培种 LD 衰减距离一般在 1.5kb 左右(Remington et al., 2001),而玉米自交系中 LD 衰减距离可达到100Kb(Rafalski et al.,2002)。除了交配系统,瓶颈效应也是 LD 的重要影响因素。作物驯化起始过程中,只有一小部分个体被选择,基因型频率收到了很大的选择,只有小部分基因被保留下来,这种现象称之为瓶颈效应。这种现象可以显着增加 LD 的水平。另外研究群体的大小也是影响 LD 的重要因素,不难想象,如果群体过小,很容易使估测的 LD 过大。
1.3.4 关联分析的一般步骤:
关联分析大体上需要经过以下几个步骤:群体构建、表型鉴定和基因型检测、群体结构分析、关联分析。具体如下:群体构建时一般需要考虑两个方面:群体大小和材料多样性,一般来说群体越大,多样性越好,研究的结果也会越好,但是成本也会增加,一般先用分子标记对种质资源进行多样性筛选,挑选出代表最广泛多态性的最小群体。这样筛选出来的群体我们可以称之为核心种质。例如本实验所用的关联分析材料为黄瓜核心种质,是由从 3342 份种质中筛选出来的 115 个株系组成,代表了总遗传多态性的 77.2%(Qi et al.,2013)。
表型的鉴定与变异分析:对于植物实验,特别是材料是纯合自交系,一般采取多年多点的重复鉴定来增加表型鉴定的准确性和稳定性。另外某些复杂性状有多个指标对其进行评估,比如黄瓜果实大小,可以包括长、直径、果重等可以通过主成分分析法,减少实验数据的采集量,找出可以代表目标性状表型的主成分因子,用其多为表型数据进行关联分析。基因型的测定:随着二代测序技术的成熟,SNP 分子标记被大量的用在关联分析上,其具有以下方面的优势:(1)标记数量多、密度大,有研究表明玉米和大刍草中,编码区每 124bp 就存在一个 SNP 位点,非编码区每 31bp 就有一个 SNP 位点;(2)检测技术成熟,包括芯片技术和二代测序技术的成熟和广泛应用,使SNP标记的检测成本降低的同时也更加快捷(汪维鹏 等,2006)。
群体结构和亲缘关系分析就是对群体中的个体进行组成分析,把所有的材料分成不同的亚群,这样可以避免相同亚群内部 LD 强度偏大导致的假阳性。关联分析所用的群体一般有如下五种:a、理想群体,具有较弱的群体结构和亲缘关系;b、多家系群体,群体结构微弱但是包含多个家系;c、具有群体结构但是亲缘关系较远;d、既有群体结构又有亲缘关系;e、具有高度群体结构和紧密亲缘关系的群体。一般来说做植物,特别是农作物多数属于第四种群体结构,所以必须进行群体结构和亲缘关系的分析。进行群体结构和亲缘关系分析需要用到覆盖全基因组的分子标记,常见的如 SSR、SNP 等,一个 SSR 分子标记常有多种基因型,而数量较少;每个 SNP位点一般只有两个基因型,但是数量巨大。所以如果是等量的分子标记的话,SSR 标记检测的效率更高(Zhu et al.,2008)表型与基因型的关联分析:Tassel 是关联分析常用的软件,关联分析所用的模型主要有 GLM(一般线性模型)和 MLM(混合线性模型)。GLM 模型:y=markereffect + populationstructure +residual。在 Tassel 软件中的 GLM 程序中将各个个体的 Q 值作为协变量,对标记分别与性状进行回归分析,计算量较少,运行时间短。MLM 模型需要先计算每个品系归属于各个亚群后 Q 矩阵和品系间亲缘关系 K 矩阵。用以矫正群体结构和遗传背景对检测结果的影响,运行时间较长。
1.3.5 关联分析在黄瓜研究中的应用
关联分析已经在重要的粮食作物中取得了丰硕的成果,如下列出了在模式作物和重要粮食作物中取得的一些重要的研究进展:在拟南芥研究中的应用:Aranzana 等通过 GWAS 技术鉴定出拟南芥中控制开花时间和抗性相关的多个信号位点(涂雨辰,2013)。在玉米育种中的应用:如 2011 年通过全基因组关联分析技术确定了一些与叶片结构相关的关键基因(Laurie et al.,2004)。在大豆分子标记研究中的应用:
在大豆中关联到一些重要的位点,这些位点与一些重要的农艺性状有关,对作声生产实践具有重大的指导作用(Yang et al.,2010;Palmer et al.,2004)。在水稻重要基因定位方面的应用:通过 GWAS技术,检测到 1 个控制花期的信号位点和多个与生产性状关联的位点(Arite et al.,2009)。虽然关联分析相对于传统的基因定位技术具有更高的效率和准确性,在重要的作物中也取得了不错的成果,但是关联分析现在在黄瓜基因功能研究中的应用尚未见报道,不过随着测序技术的成熟,以及分析技术的发展,关联分析将会更加广泛的应用在基因定位和功能研究中。
1.4 本研究的目的意义与技术路线
1.4.1 研究目的与意义
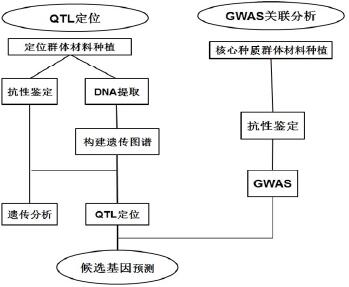
白粉病(powdery mildew)是严重影响黄瓜生产的一种病害,常造成重大的经济损失,生产上常采用过量使用杀菌剂来进行防治。大量使用药剂防治造成了生产成本增加和严重的环境污染。培育抗病新品种成为最经济、最有效的手段,所以开展对黄瓜白粉病抗性遗传规律的研究和抗病基因的克隆具有重要的实际意义。根据前人报道种质资源 PI 200815 含有抗白粉病基因 pm-3,不同于 Natsufushinari 中的 pm-1 和 pm-2 基因,也不同于控制黄瓜下胚轴抗性的 pm-h 基因。截至到目前为止,对于 PI 200815 中 pm-3 基因的定位研究尚未见报道。本研究以高抗和高感白粉病的两个材料作为抗病亲本,配制遗传群体,对抗性亲本中的抗病基因进行遗传规律分析。利用黄瓜全基因组测序开发的 SSR 分子标记,对抗病基因进行定位研究,分析候选基因,为今后分子标记辅助育种和解析抗病机制奠定基础。同时利用核心种质,通过全基因组关联分析技术,对黄瓜白粉病抗性进行全基因组关联分析,在全基因组范围内,挖掘抗白粉病基因。这样不仅可以关联到更全的基因,还可以将不同学者的研究结果进行整合分析,为综合利用黄瓜中的抗白粉病遗传资源提供依据。
1.4.2 技术路线