摘 要: 自2019年末,新冠肺炎的全球大流行使蝙蝠(翼手目动物)倍受关注,甚至人们“谈蝠色变”。蝙蝠在生态和分类上多样性极高,占哺乳动物种类的20%左右。蝙蝠不仅在害虫控制、种子传播、植物授粉等方面具有重要的生态作用和经济价值,也在健康长寿、生物仿生和语言进化等方面具有重要的科研价值。中国是全世界蝙蝠物种多样性最丰富的国家之一,已知的种类超过了140种。近几十年来,中国学者在蝙蝠分类学、生态学、行为学、进化生物学、神经生物学和病毒病源学等研究领域取得了丰硕的成果,然而涉及蝙蝠的生态系统服务和资源保护的相关研究比较少。由于生境退化、过度捕杀、栖息地破坏、城市化、杀虫剂使用、气候变化等原因,中国蝙蝠受到极大的威胁,约51%的物种位于近危等级之上。为了有效地保护中国蝙蝠物种的多样性,建议加强基础研究,建立中国蝙蝠多样性监测网络,加强蝙蝠栖息地保护,完善相关法律法规,加强蝙蝠公众教育,以促进中国蝙蝠生物学研究和保护。
关键词: 蝙蝠; 研究进展; 濒危等级; 保护;
Abstract: Since the end of 2019,the global pandemic of COVID-19 has attracted much attention on bats(Chiroptera),and even people are frequently frowned upon when talking about bats.Bats are extremely perse in ecology and taxonomy,accounting for ~20% of mammalian persity worldwide.Bats play an important role in pest control,seed transmission,plant pollination,offering a great economic benefit to human society.Moreover,bats bear important scientific research value in health and longevity,biomimetic,language evolution,and more.There are more than 140 species of bats in China,ranking China one of the top countries in terms of bat species persity of the world.In recent decades,Chinese researchers have made great progresses in perse fields such as bat taxonomy,ecology,ethology,evolutionary biology,neurobiology,and viral pathogens.However,researches focusing on ecosystem services and conservation of bats were very scarce.Due to logging and plants harvesting,over-hunting,habitat destruction,urbanization,agricultural activities,climate change,and so forth,bats in China are largely threatened:51% of bat species have been listed as Near Threatened or above(Critically Endangered,Endangered,Vulnerable).We argue that successful bat conservation in China must first strengthen basic research,particularly in assessing the population status and trends of bat species and establishing a monitoring network for bat persity.Additionally,a combination of conservation efforts,including the protection of bat habitats,construction of artificial bat habitats,improvement of relevant laws and regulations,and enhancement of public education on bats,are vital to ensuring sustainable bat populations in China.
Keyword: Bats; Research progress; Endangered categories; Conservation;

人类活动引发了全球生物多样性丧失危机(Pimm et al.,2014)。全球脊椎动物的20%已经被世界自然保护联盟(IUCN)定级为近危物种(Hoffmann et al.,2010;IUCN,2020),其中哺乳动物也不例外(Hoffmann et al.,2011)。蝙蝠是最广泛分布的陆生哺乳动物,大约1 400种,占哺乳动物的20%以上,是进化上最为成功的类群之一(Wilson and Mittermeier,2019)。其在害虫控制、种子传播、植物授粉等方面具有重要的生态作用和经济价值(Kunz et al.,2011)。在中国传统文化中,蝙蝠象征福气、长寿、吉祥、幸福(草林娣,2007)。然而,“非典”和新冠肺炎等传染病致使公众产生的恐慌,以及人们对蝙蝠缺乏系统的了解,可能导致人们对蝙蝠产生误解,从而使其遭受更大威胁,例如驱赶和捕杀(Zhao,2020)。同时,蝙蝠对环境变化极为敏感,其物种多样性更容易受到威胁,全球和中国分别约22% 和51%的蝙蝠物种位于近危等级之上(Voigt and Kingston,2016;蒋志刚等,2016)。Feijó等(2019)统计了中国2000—2017年间发表的594篇与蝙蝠相关的文章,发现保护相关的研究仅占1.6%,表明中国蝙蝠多样性保护研究和实践没有受到重视。基于这样的事实,我们有义务和责任让人们全面了解中国蝙蝠生物学研究进展、受威胁状况及保护对策,从而理性地对待蝙蝠,促进中国蝙蝠物种多样性保护。
本文概述了中国蝙蝠生物学相关领域的研究进展,分析了中国蝙蝠的受胁状况,并提出相应的物种多样性保护对策,旨在激发研究者对蝙蝠这一神秘类群的兴趣,进而加强蝙蝠的研究和保护。
1、 蝙蝠的生态学价值
1.1、 蝙蝠的生态和经济价值
蝙蝠在害虫控制、种子传播、植物授粉等方面发挥着举足轻重的作用。尽管蝙蝠食性多样,但70%的蝙蝠专性或兼性以昆虫为食(Kunz et al.,2011)。据估计,圈养的蝙蝠每天消耗的昆虫约占其体重的25%(Brisbin,1966);但在野外条件和哺乳期等高能耗时期,这个数字达70%(Kunz et al.,1995),有时甚至超过100%(Kunz and Stern,1995)。
蝙蝠经常在农田捕食多种潜在的农业害虫(Park,2015;Heim et al.,2017)。研究表明,墨西哥游离尾蝠(Tadarida brasiliensis)会伺机捕食多种农业害虫(Marais et al.,1980;Mccracken et al.,2008,2018)。Cleveland等(2006)证实墨西哥游离尾蝠每年在德克萨斯州中南部通过捕食害虫而避免棉花受损以及避免使用杀虫剂的价值为741 000美元,占棉花总产值的15%。仅在北美地区,蝙蝠通过减少作物损害和避免使用杀虫剂而产生的价值每年约为229亿美元(Boyles et al.,2011)。在泰国,蝙蝠每年在稻田中通过捕食害虫可防止稻米损失近2 900 t,其经济价值超过120万美元,意味着每年能为近26 200人提供口粮(Wanger et al.,2014)。保守估计在全球范围内,仅在玉米种植中,食虫蝙蝠通过对害虫的抑制产生的价值超过10亿美元,而蝙蝠可通过间接抑制与虫害相关的真菌和玉米上的有毒化合物而进一步造福人类(Maine and Boyles,2015)。许多农业害虫的幼虫对作物造成损害,蝙蝠捕食其成虫,从而阻止了成虫产卵,进而减少幼虫的数量,最终减少幼虫对作物的损害。因此,蝙蝠对害虫的捕食可能对农业生态系统产生级联效应(Karp and Daily,2014)。
现代分子技术的发展,例如DNA宏条形码(DNA metabarcoding)和环境DNA分析(environmental DNA analyses,eDNA)等技术,使人们对蝙蝠捕食害虫的农业服务有了新的认识。Aizpurua等(2018)通过eDNA技术对欧洲普通长翼蝠(Miniopterus schreibersii)的食性进行了分析,发现普通长翼蝠能捕食超过200种节肢动物,其中包括44种涉及欧洲大陆许多作物的农业害虫。普通长翼蝠也可根据当地农田中可利用食物资源调整食性,重塑其食性生态位(Aizpurua et al.,2018)。因此,以往蝙蝠对农业害虫的抑制作用被严重低估。
蝙蝠还可以授粉和传播植物种子,提供关键的生态系统服务功能(Bumrungsri et al.,2008,2009)。研究表明,虽然大蜜蜂(Apis dorsata)是榴莲最频繁的访花动物,但果蝠,尤其是大长舌果蝠(Eonycteris spelaea)是榴莲的主要传粉者(Bumrungsri et al.,2009)。在热带地区,狐蝠是高效的传粉者和种子传播者,约23%的植物依赖其传播种子(Fujita et al.,1991;Roberts et al.,2012)。在非洲,大约34%经济木材的种子需要依靠狐蝠进行扩散(Muscarella and Fleming,2007)。在岛屿,进化的偶然性和人为导致的其他种子传播者的灭绝,致使狐蝠成为授粉或传播种子的唯一媒介,进而成为岛屿上维持植物生存的关键种(Mcconkey and Drake,2015)。因此,岛屿上狐蝠的灭绝可能会引发连锁灭绝反应(Pierson et al.,1996),从而导致不可挽回的生态和经济后果。
1.2、 蝙蝠的科研价值
蝙蝠因其免疫功能强大、患癌概率极低和超长的寿命成为人类医学绝佳的研究对象(Foley et al.,2018;Seluanov et al.,2018)。首先,蝙蝠是携带人类烈性病毒的近缘种类,例如新冠病毒(SARS-CoV-2)、严重急性呼吸综合征病毒(SARS-CoV)、埃博拉病毒(Ebola virus)等。令人惊奇的是,与人类及其他哺乳动物不同,携带致命病毒的蝙蝠却不会表现出明显的临床症状(Banerjee et al.,2020)。研究表明,蝙蝠天然免疫中的一些组分相较于其他哺乳动物更为活跃,使其免疫系统始终处于警惕状态,有效地抑制病毒复制(Xie et al.,2018)。另一方面,蝙蝠体内许多与过度免疫和炎症反应相关的分子却在表达和功能上受到抑制,避免组织器官在抗病毒期间受到损伤(Ahn et al.,2019)。因此,对蝙蝠免疫机制的研究可以帮助人类更好地理解疾病的发生与控制,探究对抗病毒的新手段,进而开发出新的治疗方式。其次,一项对亚洲、非洲和澳大利亚蝙蝠的病理学研究表明,蝙蝠患癌概率极低(Seluanov et al.,2018)。一个编码转运体的基因ABCB1会在蝙蝠体内高表达,显着抑制蝙蝠细胞的DNA损伤,从而降低蝙蝠的肿瘤发生率(Koh et al.,2019)。因此,深入研究蝙蝠的抗癌机理,有助于加深和扩展人们对致癌机理认识,进而研发抑制或治愈癌症的药物和工具。最后,虽然蝙蝠体型较小,但寿命极长,有些蝙蝠的寿命可达40年之久,约为相似体型哺乳动物寿命的8倍(Lagunas-Rangel,2020)。如果能像蝙蝠一样长寿,按体积换算,人类可活240年。研究发现,与其他哺乳动物不同,长寿蝙蝠的端粒不会随着年龄的增加而缩短,并且与DNA修复相关的基因和抑癌基因在长寿蝙蝠中经历了强烈的适应性选择(Foley et al.,2018)。因此,加强蝙蝠长寿机制的研究,能为人类实现健康长寿的目标提供新的理论基础。
蝙蝠独特的回声定位能力和高超的飞行技巧成为生物仿生良好的研究对象(张树义,2009)。研究表明,蝙蝠回声定位已在自然选择压力下不断完善并使用了约6 500万年(Teeling et al.,2005)。因此,蝙蝠的回声定位具有智能的认知能力、超强的抗干扰本领、快速而精准的适应性调控(Jakobsen et al.,2015),能为解决现代雷达面临的难题提供新的视角和手段。此外,蝙蝠是唯一真正会飞的哺乳动物,有着令人难以置信的灵巧与机动性。其飞行堪称是世界上最离奇、最完美的运动,鸟类和昆虫都无法与之媲美(Hedenstrom et al.,2007)。因此,深入研究蝙蝠的飞行技巧,并且应用到现代飞行器上,有助于实现“智能飞行”和“安全飞行”。
蝙蝠是少数具备发声学习能力的哺乳动物之一。与多数哺乳动物不同,回声定位蝙蝠具有极高的发声活跃性(每秒发声数量从几个到上百个不等),为研究发声控制的脑机制提供了便利的行为模式。蝙蝠在研究声音通讯和导航方面具有独特优势,对于蝙蝠发声控制和学习的研究使其成为动物声音通讯研究的主要模式物种之一,也成为破解人类语言脑机制的希望(Luo et al.,2018)。此外,有些蝙蝠能够迁徙,每晚需要飞到数十公里外捕食,它们如何实现如此大尺度的空间导航?对其进行研究,有助于揭示人和动物大脑方位感知和空间导航的秘密。
2、 中国蝙蝠物种多样性及分类学
2.1、 新种与新纪录发表
查询到有关我国蝙蝠区系分类与分布研究相关论文210篇,其中2000年以来共发表175篇,占83.3%,年均8.75篇,约为之前20年年均篇数的5倍多(表1)。4种类型的论文数量都迅猛增长,尤其是发表新种论文达到13篇(合计新种17种),新纪录(包括中国新纪录、省级新纪录等)报道多达116篇,占同期论文总数的55.2%。
表1 我国(大陆)蝙蝠区系分类与分布期刊论文的分布情况
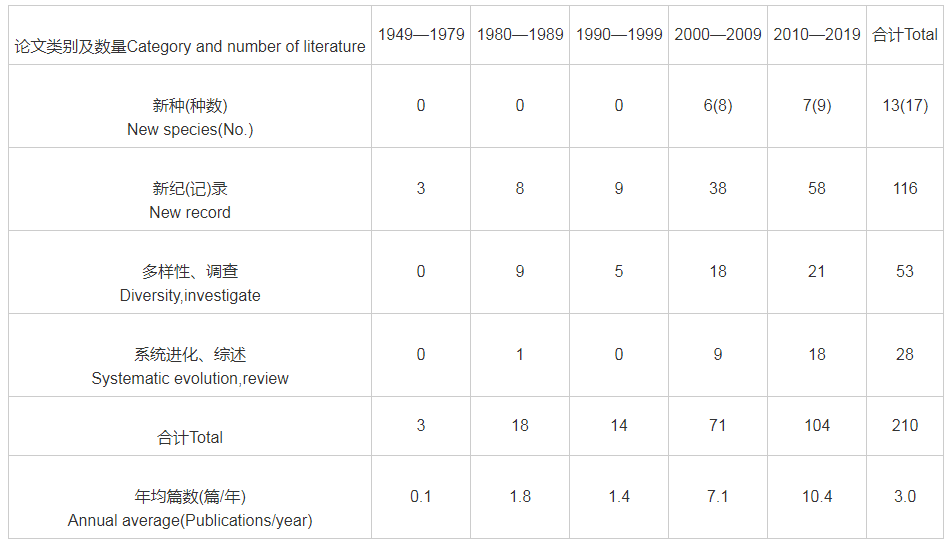
从18世纪中期开始至新中国成立,我国的蝙蝠研究主要由外国人开展。自我国台湾学者Kuo等(2006)发表金芒管鼻蝠(Harpiola isodon)新种之后,蝙蝠研究学者不断探索,至2019年底,先后发表了中国境内的蝙蝠新种共计17个,管鼻蝠属新种多达10个(表2)。但部分种类有效性存在一定争议,如有学者认为华南菊头蝠(Rhinolophus huananus)(Wu et al.,2008)应该是清迈菊头蝠(Rhinolophus siamensis)(Zhang et al.,2009;Zhang et al.,2018);榕江管鼻蝠(Murina rongjiangensis)(Chen et al.,2017)似乎与水甫管鼻蝠(Murina shuipuensis)(Eger and Lim,2011)外部形态相近,亲缘关系极近;倭扁颅蝠(Tylonycteris pygmaeus)(Feng et al.,2008)和北京宽耳蝠(Barbastella beijingensis)(Zhang et al.,2007)等新种有效性也有待进一步确认。
从中国蝙蝠新纪录方面看,除2000年之前发表的大狐蝠(Pteropus giganteus)(王宗禕和汪松,1962)、缺齿鼠耳蝠(Myotis annectans)(罗一宁,1987)、泰国狐蝠(Pteropus lylei)(何晓瑞和杨白仑,1991)3个种之外,进入21世纪后陆续发现了高鞍菊头蝠(Rhinolophus paradoxolophus)(赵辉华等,2002)、小巨足鼠耳蝠(Myotis hasseltii)(张礼标等,2004)、马氏菊头蝠(Rhinolophus marshalli)(吴毅等,2004a)、小褐菊头蝠(Rhinolophus stheno)(张劲硕等,2005)、泰国无尾果蝠(Megaerops niphanae)和无尾果蝠(Megaerops ecaudatus)(冯庆等,2006)、安氏长舌果蝠(Macroglossus sobrinus)(冯庆等, 2007)、大趾鼠耳蝠(Myotis macrodactylus)(江廷磊等,2008)、灰小蹄蝠(Hipposideros cineraceus)(谭敏等,2009)、艾氏管鼻蝠(Murina eleryi)(刘志霄等,2014)、马来假吸血蝠(Megaderma spasma)(张礼标等,2010)、哈氏管鼻蝠(Murina harrisoni)(吴毅等,2017)、泰坦尼亚彩蝠(Kerivoula titania)(Wu et al.,2012)、印支鼠耳蝠(Myotis indochineses)(Wang et al.,2017)、环颈伏翼(Thainycteris aureocollaris)(Guo et al.,2017)等中国蝙蝠新纪录15种。省级蝙蝠新纪录发现较多,共发表论文101篇(占同期论文的 48.1%)。
表2 2006年以来中国境内发表的翼手目新种
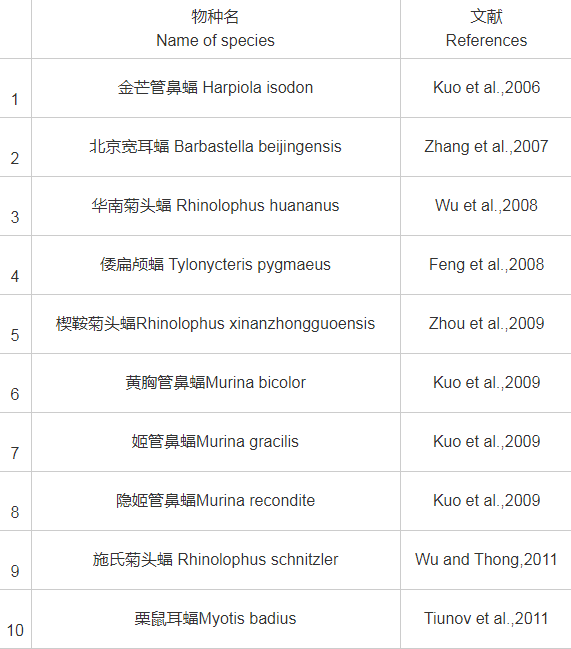
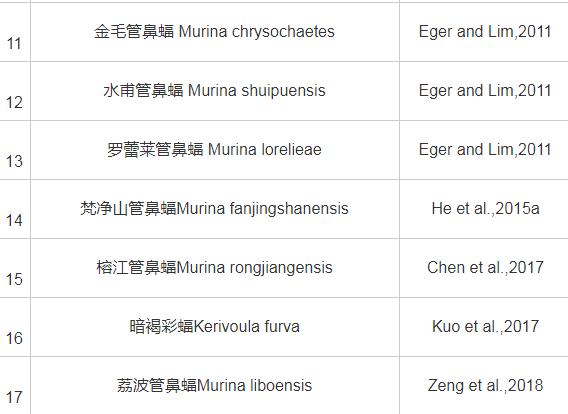
2.2 、分类学问题探讨
由于蝙蝠飞行和夜行生活的特殊性,国内外对其分类学研究均相对滞后。随着调查强度的加强与捕捉方法的更新(如竖琴网的使用),尤其是分子系统学证据、回声定位超声波、染色体与多元统计分析技术的综合应用,使传统形态学研究方法不易区分的物种有了被区别和鉴别的可能。吴毅等(2004b)依据核型,将我国分布的鲁氏菊头蝠中华亚种(Rhinolophus rouxi sinicus)提升为种,即中华菊头蝠(Rhinolophus sinicus);Li 等(2006)从分子角度、Wu 等(2012)从数值分类角度对东亚分布的小菊头蝠(Rhinolophus pusillus,中国大陆)、单角菊头蝠(Rhinolophus monoceros,中国台湾)、角菊头蝠(Rhinolophus cornutus,日本本岛)及角菊头蝠琉球亚种(Rhinolophus c.pumilus,日本冲绳)进行了比较,前者将它们视为不同的地理亚种,而后者认为形态有一定差异,仍应独立为不同种;Sun 等(2008)在江西发现了菊头蝠的一个隐蔽种,该结果也与Wu 等(2011)在广东英德(模式产地)发现的华南菊头蝠(Rhinolophus huananus)新种相吻合;Huang 等(2014)从分子、染色体和超声波等对华南分布的扁颅蝠(Tylonycteris pachypus)进行了比较,建议将该种的华南亚种(Tylonycteris p.fulvidus)提升为种,定名为华南扁颅蝠(Tylonycteris fulvida),已得到国际同行的认可(Wilson and Mittermeier,2019);党飞红等(2017)对原广泛分布于中国的绯鼠耳蝠(Myotis formosus)进行了学名厘定,确定耳廓端部和拇指为红褐色者应为渡濑氏鼠耳蝠(Myotis rufoniger),而无上述特征,且全身为纯浅黄色者为金黄鼠耳蝠(Myotis formosus)(主要分布在台湾及江西);Yu 等(2018)对原认为中国广泛分布的哈氏彩蝠(Kerivoula hardwickii)及泰坦利亚彩蝠(Kerivoula titania)进行了比较,确定应该订正为华南彩蝠(Kerivoula furva)。
我国先后出版寿振黄(1958,1962)、夏武平(1964)、钱燕文等(1965)编写的兽类学专着及部分地方兽类志(或动物志)。21 世纪以来,主要兽类专着有《中国哺乳动物物种和亚种分类名录与分布大全》(王应祥,2003)、《中国哺乳动物多样性和地理分布》(蒋志刚,2005)、《中国哺乳动物彩色图鉴》(潘清华等,2007)、《中国兽类野外手册》(Smith和解焱,2009)和《中国兽类图鉴》(刘少英和吴毅,2019)等。在这些专着及地方兽类志中,研究者系统地澄清了中国蝙蝠分类学的很多问题。
3、 蝙蝠生物学研究进展
3.1、 蝙蝠种群生态学
蝙蝠在中国超过140种,占全球蝙蝠物种约10%,物种多样性极高。蒋志刚等(2016)依据IUCN的标准,对中国134个蝙蝠物种划分濒危等级。结果表明,3种蝙蝠濒危(Endangered,EN,2.24%)、15种蝙蝠易危(Vulnerable,VU,11.20%)、51种蝙蝠近危(Near Threatened,NT,38.06%)、42种蝙蝠无危(Least Concern,LC,31.34%)、23种蝙蝠数据缺乏(Data Deficient,DD,17.16%)。尽管琉球狐蝠(Pteropus dasyma-llus)、安氏长舌果蝠(Macroglossus sobrinus)和彩蝠(Kerivoula picta)被列为濒危物种(蒋志刚等,2016),但并没有针对性的物种恢复计划和措施。数据显示中国51%的蝙蝠物种位于近危及其之上等级,而全球仅22%的蝙蝠物种位于近危及其之上等级(Voigt and Kingston,2016)。这表明中国的蝙蝠受到更大程度的威胁,且中国缺乏蝙蝠物种多样性保护的研究和实践。然而,迄今为止,尚没有任何一种蝙蝠物种列入最新的“中国国家重点保护野生动物名录”。
种群数量的动态监测,是物种多样性保护计划制定和实施的前提。根据我们近20年的野外调查,中国的蝙蝠种群数量与2000年相比可能下降超过50%(Zhao,2020)。然而,在中国,长期的蝙蝠物种多样性和种群数量监测的研究相对较少,仅涉及少数地区和物种。例如广州地区扁颅蝠(Tylonycteris pachypus)的种群数量与相对湿度显着负相关(吴毅等,2004),而东北地区的马铁菊头蝠(Rhinolophus ferrumequinum)秋季飞行活动与温、湿度显着正相关(Jiang et al.,2016);扁颅蝠和褐扁颅蝠(Tylonycteris robustula)集群大小分别为1~24只和1~13只(张礼标等,2004);人为干扰显着影响大卫鼠耳蝠(Myotis davidii)和大蹄蝠的集群数量(Luo et al.,2012;由玉岩,2013)等。此外,河南省常见的蝙蝠物种(刘伟等,2011),尤其是马铁菊头蝠(Niu et al.,2007),以及东北地区蝙蝠物种多样性及相对数量已经基本调查清楚(Bat Ecology,Evolution and Conservation Lab,2019)。
中国蝙蝠生态学研究,最早可追溯到1959年。着名动物生态学家熊郁良先生1959—1965年对昆明花红洞的9种蝙蝠的昼夜活动规律、数量变动、种群结构、繁殖及食性等进行了长期观察(熊郁良,1975)。之后,中国蝙蝠生态学研究迅速发展,其主要集中在蝙蝠的食性(Zhang et al.,2005;Ma et al.,2006a,2010)、捕食策略(金龙如等,2005;Jin et al.,2005)、生境选择(Wang et al.,2010;Bu et al.,2015;龚小燕等,2018;Liu et al.,2018)、体型变化(Jiang et al.,2019b)、分布(刘伟等,2011;胡宜峰等,2019;Luo et al.,2019)和种间关系(罗峰等,2005;Jiang et al.,2008;王静等,2010;Jiang et al.,2013)等方面。这些表明中国蝙蝠生态学研究的科学问题逐渐深入,研究方法也日趋先进。例如食性研究从单纯的描述到食性生态位的适应和进化机制(Chang et al.,2019;Shi et al.,2020),研究方法也从传统的粪便分析法转变到高通量测序技术(Chang et al.,2019)。然而,与蝙蝠进化生物学和声行为相比,蝙蝠生态学方面的研究还十分欠缺。因此,将来应结合先进的技术手段,例如高通量测序、热红外成像、声学监测和微型GPS等技术,来解决经典的蝙蝠生态学问题。
3.2 、蝙蝠声行为
中国蝙蝠声行为研究在2000年后才系统地开展。2000—2019年,中国蝙蝠声行为相关的研究成果在数量和质量上都呈上升趋势。在回声定位行为方面, 从早期的主要集中于对一种或几种蝙蝠回声定位声波特征进行简单描述,拓展至回声定位声波特征和功能(Wang et al.,2010)、声波与形态特征的关系(Feng et al.,2002,2012)、回声定位声波的可塑性与地理进化(Jiang et al.,2015;Luo et al.,2019)以及通过回声定位声波进行物种鉴定等(Wu et al.,2020)。国内的蝙蝠交流声波研究开始于2013年后,主要围绕交流声波特征、多样性、发声背景与功能及可塑性开展工作(Lin et al.,2015;Guo et al.,2019;Zhao et al.,2019)。
近年来,关于蝙蝠回声定位声波可塑性及地理进化的深入研究不断增多。在多种蝙蝠中发现不同种群的回声定位声波具有显着的地理差异,其进化驱动力可能与生态选择、文化漂变等因素有关(Zhang et al.,2019;Song et al.,2019)。而且,蝙蝠能够根据背景噪声对回声定位声波的发声频率、振幅等进行调节,以避免噪声干扰(Luo et al.,2019)。与回声定位声波相比,交流声波音节类型复杂而多样(Ma et al.,2006b),且在捕食、激进、胁迫等不同行为背景下使用的音节类型不同,并且具有一定的句法结构(Lin et al.,2015;Guo et al.,2019)。因此,蝙蝠交流声波在个体竞争与社群稳定性维持中起重要的作用(Luo et al.,2017;Xiao et al.,2018;Jiang et al.,2019;Sun et al.,2019;Zhang et al.,2019;Zhao et al.,2019)。此外,幼蝠发育过程中的隔离叫声(isolated calls)随年龄而变化,而且具有母婴识别的功能(Jin et al.,2015;Chi et al.,2019)。这些研究表明我国蝙蝠声行为研究正在向揭示动物声信号功能、形成与进化机制及探索与人类语言进化相关的科学问题深入。
综上所述,中国蝙蝠声行为研究虽然起步较晚,但是近年来发展迅速,研究内容也在不断深入和多元化。但是,研究水平仍需进一步提升,尤其在回声定位行为方面。国际上的前沿研究已集中于使用麦克风阵列技术研究蝙蝠声场特征与3D导航机制(Wohlgemuth et al.,2018),而我国目前的研究还多使用单一麦克风录制和分析二维声谱参数。在社群交流叫声方面,还需开展更多的关于交流声波进化驱动力、发声学习等研究。未来的蝙蝠声行为研究应采用更先进的实验设备与技术手段来解决更深层的发声机制与进化问题。
3.3、 蝙蝠进化生物学
3.3.1、 回声定位的进化机制
FoxP2基因与人类的语言发育相关。研究发现蝙蝠的FoxP2基因序列变化很大,这与FoxP2在其他非人哺乳动物中的高度保守截然不同,提示FoxP2的加速进化可能与蝙蝠独特的发声机制有关(Li et al.,2007)。 Li 等(2008)发现听觉相关基因Prestin在回声定位蝙蝠中发生了趋同进化,揭示Prestin的功能可能与蝙蝠回声定位的能力密切相关,随后,发现Prestin在齿鲸(toothed whales,Odontoceti)和蝙蝠中趋同进化,表明Prestin在回声定位物种中经历了相似的选择压力,证实了Prestin的功能与回声定位能力的起源与演化密切相关(Li et al.,2010;Liu et al.,2010)。然而,以上研究都是基于序列分析的结果,缺少实验验证。Liu等(2014)使用膜片钳技术对Prestin进行细胞实验,结果显示回声定位物种Prestin基因在序列水平上的趋同会导致功能上的趋同。Li 等(2016)分析了具备不同回声定位能力蝙蝠的Prestin基因序列,结合体外细胞实验,鉴定出一个与蝙蝠调频回声定位能力相关的位点,被认为是蝙蝠祖先的多态位点。此外,一些听觉相关基因可能也与蝙蝠独特的回声定位能力有关(Liu et al.,2011,2012a;Shen et al.,2012)。随着基因组学的快速发展,基于组学数据来揭示蝙蝠回声定位能力的分子机制也开始涌现。对两种恒频蝙蝠进行全基因组测序,鉴定出了一些未报道过的、可能与蝙蝠恒频回声定位相关的基因(Dong et al.,2016);基于蝙蝠转录组数据也鉴定出一些新的与蝙蝠回声定位能力有关的基因(Wang et al.,2019)。
3.3.2、 视觉相关基因
Zhao等(2009)发现蝙蝠的中/长波视蛋白基因M/LWS在物种间非常保守,而短波视蛋白基因SWS1在一些具有恒频回声定位能力的旧大陆蝙蝠物种和洞穴生活的蝙蝠物种中发生了假基因化,这可能与恒频回声定位能力的起源以及蝙蝠栖息环境的改变有关。SWS1基因在独立起源的新大陆恒频蝙蝠中也发生了假基因化,证实了SWS1的假基因化与恒频回声定位能力的起源密切相关,被认为是感觉代偿的一种方式(Wu et al.,2018)。研究还发现,3种吸血蝙蝠的SWS1都发生了假基因化,这可能与其特化的红外感知能力发生了感觉代偿(Li et al.,2018;Wu et al.,2018)。此外,Shen等(2010)对蝙蝠的视紫红质基因RH1进行了研究,鉴定出了多次趋同演化事件,揭示了RH1的分子进化与蝙蝠的生境(例如光环境)有关。
3.3.3 、食性相关基因
蝙蝠是哺乳动物目级分类阶元中最大的食性适应性辐射,多次独立演化出食果、食蜜、食肉和杂食等食性(Jiao et al.,2019)。Zhou等(2009)首次发现莹鼠耳蝠(Myotis lucifugus)的部分苦味受体基因受到了正选择作用,揭示苦味受体的功能对食虫蝙蝠至关重要。吸血蝙蝠的苦味受体基因很多都发生了假基因化,其比例显着地高于其他食性蝙蝠,证实吸血蝙蝠的苦味觉退化,可能与其专一的食性有关(Hong and Zhao,2014)。最新的研究使用基因组数据鉴定了15种蝙蝠的苦味受体基因家族,发现食果蝙蝠和食虫蝙蝠的苦味受体基因的数目并没有显着差异(Jiao et al.,2018)。该研究发现鼠耳蝠属特异的基因复制事件,使得鼠耳蝠的苦味受体基因数目明显多于其他蝙蝠,且复制出的苦味受体能感知新的苦味物质,表明其能够帮助鼠耳蝠更快地适应新的环境,可能是鼠耳蝠属发生适应辐射的分子基础之一(Jiao et al.,2018)。此外,蝙蝠的甜味受体基因Tas1r2在大部分蝙蝠中保守,但在3种吸血蝙蝠中都变成了假基因,首次从分子生物学的角度验证了行为学实验的结果(Zhao et al.,2010)。鲜味受体基因Tas1r2在所研究的蝙蝠中都发生了假基因化或者丢失,表明鲜味觉对蝙蝠并不重要,但不需要鲜味感知的原因仍需进一步研究(Zhao et al.,2012)。
Liu等(2012b)对蝙蝠的丙氨酸乙醛酸盐转氨酶(AGT)基因进行了研究,发现其线粒体靶向序列在部分食果蝙蝠中发生了丢失,导致AGT不能定位到线粒体,这可能与蝙蝠食性的转变相关。研究发现食虫蝙蝠的C型溶菌酶基因(Lyz)发生了复制,推测食虫蝙蝠复制出来的Lyz基因能够提高其对昆虫外壳几丁质的降解能力(Liu et al.,2014)。最近,研究者发现所有的食虫蝙蝠都具有完整海藻糖酶基因(Treh),而许多非食虫蝙蝠的变成了假基因,表明Treh基因对于食虫蝙蝠不可或缺(Jiao et al.,2019)。
3.4、 蝙蝠神经生物学
30多年来,我国研究者在蝙蝠听觉神经生物学领域取得了一定的成绩。其中,研究涉及到蝙蝠听觉系统的发育可塑性(孙心德等,1992;周晓明和孙心德,1997)、蝙蝠听觉处理的空间差异性(Sun and Jen,1986;Zhang et al.,2000)、不同脑区生理学活动对蝙蝠听觉信息处理的影响(Sun and Jen,1989;Zhang et al.,2001)、蝙蝠听觉可塑性的分子机制(周晓明和孙心德,1999;徐丽静和孙心德,1998)、蝙蝠听觉系统对不同回声定位声波参数的编码(Luo et al.,2008;Yang et al.,2018),声波参数动态变化对信息编码的影响(Fu et al.,2008;Tang et al.,2017)和不同脑区生理学活动对蝙蝠听觉信息处理的影响(Peng et al.,2017)等内容。研究的技术手段则涵盖了电生理学(胞内记录、胞外记录和膜片钳)、神经形态学、生物化学等。最近,昆明动物所张亚平院士团队结合电生理学、核磁共振成像、分子生物学等手段对蝙蝠的暗视觉开展比较研究(Liu et al.,2015),反映出我国蝙蝠神经生物学的研究已经进入了学科交叉的新时代。
3.5 、蝙蝠病毒学
蝙蝠是哺乳动物病毒最重要自然宿主之一(Jones et al.,2008;Moratelli and Calisher,2015),目前有40个科的病毒能感染脊椎动物,而从蝙蝠中发现的病毒就有28个科(Moratelli and Calisher,2015)。2003年SARS(Severe acute respiratory syndrome coronavirus,SARS CoV)以后,促使了野生动物病毒生态学的研究,蝙蝠病毒的研究在我国也进入了一个持续高热的时期,随之而来的是越来越多的蝙蝠病毒被发现,其中不乏一些具有重要公共卫生或者生物学意义的病毒,比如狂犬病病毒(Liu et al.,2013)、冠状病毒(Lau et al.,2005;Li et al.,2005)、丝状病毒(He et al.,2015b;Yang et al.,2019)、汉坦病毒(Guo et al.,2013;Xu et al.,2019)等,其中蝙蝠冠状病毒(Coronavirus,CoV)备受关注。
目前,冠状病毒有38种,其中16种与蝙蝠相关。2002年末暴发的SARS其病原就追溯到了中国南方的菊头蝠(Lau et al.,2005;Li et al.,2005);同时我国蝙蝠中还发现了很多其他SARS CoV的变异株(Ge et al.,2013;He et al.,2014;Hu et al.,2017),有力地证实蝙蝠是SARS CoV的自然宿主。2012年,暴发于沙特阿拉伯的中东呼吸综合征(Middle East respiratory syndrome,MERS)是第二个引发全球重视的冠状病毒病,MERS CoV与我国蝙蝠冠状病毒HKU4和HKU5在基因水平上高度相关(Wang et al.,2014;Yang et al.,2014)。2016年底,广东省猪场暴发了不明原因的急性腹泻,仔猪死亡率高达90%,是由一种与蝙蝠冠状病毒HKU2有95%的同源病毒(SADS CoV)引起的(Zhou et al.,2018)。2019年末暴发的新冠肺炎(COVID-19)是由一种新型SARSr CoV(即SARS-CoV-2)引起,该病毒与SARS CoV的全基因组水平同源性为79%,与浙江省某菊头蝠SARSr CoV ZC45的同源性高达89%(Wu et al.,2020),而与云南的中菊头蝠(Rhinolophus affinis)SARSr CoV RaTG13的同源性高达96%(Zhou et al.,2020),因此,SARS-CoV-2很可能是由蝙蝠SARSr CoV演变而来。鉴于蝙蝠冠状病毒具有如此丰富的多样性,同时不同冠状病毒之间容易重组,进而产生新病毒,因此蝙蝠冠状病毒在未来依然是监测的重点。
4、 蝙蝠面临的主要威胁
虽然蝙蝠在自然界具有极高物种多样性,但目前超过一半蝙蝠物种的种群数量都在急剧下降(Frick et al.,2019),其中,我国蝙蝠种群数量在20年内下降了50%(Zhao,2020)。保护工作有效开展的重要前提是确定蝙蝠种群所面临的主要威胁。目前,中国蝙蝠面临的主要威胁包括以下几个主要方面。
4.1 、生境退化及栖息地破坏
采伐是对森林最常见的干扰之一,导致森林严重退化(简耘等,2008)。Clarke 等(2005)发现森林砍伐造成大量原始森林丧失,蝙蝠的食物短缺,迫使蝙蝠外迁,与人为邻的概率增加。此外,森林砍伐导致大量树栖蝙蝠丢失栖息地,迫使蝙蝠无家可归,种群数量急剧下降(Andrews,2018)。在中国建国初期,由于基础设施建设和钢铁炼制,对木材的需求量非常大,导致我国森林资源大幅度减少(刘元,2015)。虽然没有相关研究,但中国森林资源的减少可能对蝙蝠物种的种群数量产生了负面的影响。
蝙蝠的栖息地多种多样,包括天然山洞、防空洞、人工建筑物(如房屋、桥梁等)、树叶、树洞等。栖息地对蝙蝠生存至关重要(Fenton,2001),栖息地破坏必将对蝙蝠种群繁衍造成严重的影响。岩溶洞穴旅游开发对洞穴生态环境产生显着影响,包括水量减少、温度升高、湿度降低、CO2浓度增加、空气负离子浓度减小以及洞中蝙蝠、斑灶马、蜘蛛数量减少等(邓亚东等,2011)。人类在洞穴内的活动,或者开山过程中对洞穴的直接破坏,均对洞内蝙蝠产生影响,造成蝙蝠生存力下降或弃洞。此外,中国改革开放后,城市化步伐加快(汪冬梅,2003),古式建筑物逐渐被现代化的钢筋水泥替代,原来那些逐渐适应了将人类建筑物作为栖息地的蝙蝠面临栖息地大量减少的窘境。
除此之外,滥捕滥杀对蝙蝠的生存也构成一定程度的影响(Zhang et al.,2009)。2002—2003年暴发的SARS疫情,病毒源头证实来源于蝙蝠(Li et al.,2005;Ge et al.,2013),此后对蝙蝠的直接食用大大减少。始于2019年底的COVID-19疫情,目前科学家仍在努力开展溯源研究,迄今为止发现最接近的病毒也来自蝙蝠(Zhou et al.,2020)。国家层面在野生动物禁食方面进行了专门的立法,对蝙蝠的保护带来极大的好处。
4.2、 城市化
蝙蝠占据夜空生态位,对光污染极为敏感(Stone et al.,2015)。虽然灯光能够吸引大量昆虫在光源聚集,为伏翼、棕蝠和山蝠等提供觅食机遇(Haddock et al.,2019),但人造灯光对蝙蝠的出飞、栖息、繁殖、通讯和捕食行为都产生一定的负面影响(Stone et al.,2015)。中国自改革开放以来,城市化发展迅速,光污染日益加剧。然而,迄今为止中国尚无关于人造灯光对蝙蝠影响的研究。但光污染已经对中国蝙蝠产生了极大的影响。
相对于光污染,人为噪声对中国蝙蝠的影响已经引起了学者们的关注。研究发现,环境噪声显着影响了蝙蝠回声定位声波的频谱时间参数,例如声强和频率的增加(Hage et al.,2013;Luo et al.,2015a)、声音结构的改变等(Hage et al.,2014;郭熊等,2015)。环境噪声也影响了蝙蝠的捕食活动和成功率(Luo et al.,2015b;孙淙南等,2016)。除了回声定位声波,交通噪声也影响了蝙蝠通讯叫声的强度、复杂性、持续时间和发声速率,暗示蝙蝠通过调整通讯叫声来响应噪声的干扰(Jiang et al.,2019a;Song et al.,2019)。这些研究大多在实验室进行,将来应在野外开展噪声污染对蝙蝠捕食效率、繁殖成功率和种群数量等方面的研究。
4.3 、杀虫剂和化肥的使用
蝙蝠是杀虫剂、化肥、工业污染物的高危暴露群体。有机污染物对蝙蝠具有急性和慢性效应。急性效应泛指蝙蝠摄入高浓度污染物致死,或者觅食被严重污染的昆虫和果实中毒死亡。国际上关于杀虫剂致死蝙蝠已有报道(Bennett and Thies,2007)。慢性效应指当蝙蝠摄入较低浓度的有机污染物,并不立即死亡,但由于生物富集作用,呈现慢性效应,表现为DNA变异、免疫力降低、内分泌紊乱及空间导航能力受损(Bayat et al.,2014)。例如感染白鼻综合症的莹鼠耳蝠体内多氯联苯(PCBs)、多溴联苯醚(PBDEs)、 二氯二苯三氯乙烷(DDT)和氯丹的富集量均高于未感染疾病的个体(Kannan et al.,2010)。
中国目前关于农药和化肥使用与蝙蝠相关性的研究只有零星报道。中国广西的皱唇犬吻蝠(Chaerephon plicata)种群在过去的几十年中,从100万只降低到10万只左右,可能与杀虫剂的过量使用相关(Zhang et al.,2009)。Liu 等(2018)发现中国西南地区绒山蝠(Nyctalus plancyi)毛发内的重金属镉含量在1975—2016年间不断上升,这是因为在绒山蝠捕食的玉米田内大量使用含镉的鳞酸盐化肥,说明重金属镉在蝙蝠体内存在生物富集和跨生态系统(Cross-ecosystem)传递。Heiker等(2018)对中国四川成都的普通伏翼(Pipistrellus abramus)和绒山蝠进行汞富集检测,发现所有被检测蝙蝠体内的汞富集均超过与免疫力降低相关的阈值(0.2 mg/kg),而大多数普通伏翼(7/13)体内汞超过内稳态控制和影响运动能力的阈值(10 mg/kg)。由此可见,在中国,过量使用化肥和杀虫剂已对蝙蝠造成了多方面的影响(Hu et al.,2016),但是,相关研究仍十分缺乏,亟需引起足够的重视。
4.4 、全球气候变化
全球平均气温的上升导致蝙蝠分布范围的变化(LaVal,2004;Sachanowicz et al.,2006;Lundy et al.,2010;McCracken et al.,2018)、春季迁徙时间提前(Stepanian and Wainwright,2018)、生产后代时间提前(Frick et al.,2010;Stepanian and Wainwright,2018)和夜晚外出捕食开始时间改变(Frick et al.,2012)等。全球平均气温的上升也可能会通过影响昆虫猎物的数量和分布、蝙蝠授粉植物物种的开花时间、蝙蝠在冬眠过程中的能耗等对蝙蝠产生间接的影响(Sherwin et al.,2012)。也有建模的数据指出全球气温的上升可能会改变回声定位蝙蝠的猎物探测能力(Luo et al.,2014)。
近年来,越来越多的报道证实全球极端气候事件也是影响蝙蝠种群动态的关键因素(Frick et al.,2019)。例如澳大利亚发生过的19次极热事件(>42℃的高温)造成了至少3万只狐蝠的死亡(Welbergen et al.,2008)。不同区域的台风事件可能通过摧毁蝙蝠的栖息地或捕食生境导致某些蝙蝠种类高于50%的种群减少(Craig et al.,1994;McConkey et al.,2004),但某些蝙蝠的种群大小似乎不太受台风的影响(Scanlon et al.,2018)。此外,极度干旱、极度寒冷、过度降水等极端气候事件都可能导致蝙蝠种群的下降(Jones et al.,2009)。我国是世界上自然灾害最严重的几个国家之一(Fischer and Knutti,2015),但至今尚无关于全球气候变化对蝙蝠影响的研究报道。
4.5 、公众对蝙蝠的误解
蝙蝠在中国传统文化中始终象征福气、长寿和吉祥。然而,2003年的SARS、2014年的埃博拉出血热以及2019年的新冠肺炎等人类新发传染病,不断将蝙蝠推向风口浪尖,让其以“病毒库”的角色多次出现在公众视野中(Feijó et al.,2019;Zhao,2020),甚至有人开始谈“蝠”色变,建议对其展开生态捕杀。同时,受一些西方传说或电影中蝙蝠吸血鬼形象的影响,国内一些公众误认为许多蝙蝠都以吸血为生,从而对蝙蝠存在偏见。实际上,世界上仅有的3种吸血蝠仅分布美洲中部和南部,而中国的蝙蝠多以农林害虫为食。
此外,一些公众可能会担心被蝙蝠咬伤从而感染狂犬病毒,实际上,蝙蝠通常栖息在洞顶、树洞、建筑物缝隙等人类难以抵达的地方,不会主动攻击人类。依据蝙蝠世界保护区(Bat World Sanctuary)的报道,全世界每年有3万多人死于狂犬病,其中99%由狗造成,因猫、蝙蝠等导致的人类狂犬病案例尚不足总数的1%(http://h-s.batworld.org.cslg.naihes.cn/rabies-info/)。事实上,所有的野生动物都可能携带各类病毒,而人类与野生动物频繁接触无疑加速了病毒从野生动物向人类的溢出。当前,波及全球的新冠肺炎,其病毒是否来源于蝙蝠尚无定论,若公众对蝙蝠存在误解,只关注蝙蝠对人类有害的一面,而忽视蝙蝠对人类社会和生态系统的巨大贡献,势必会造成大量无辜蝙蝠被有意或无意地杀害(Zhao,2020)。
5 、对中国蝙蝠物种多样性研究和保护建议
5.1 、加强基础研究
5.1.1 、澄清蝙蝠的生态和经济价值
尽管很多研究证实了蝙蝠在生态系统中具有重要的生态作用和经济价值,然而,其研究区域主要集中在美洲、欧洲、非洲和东南亚地区,且重点关注常见的农作物及其地区性的经济作物。对于物种数量占全球10%的中国,仅见蝙蝠在生态系统中作用的零星报到(Jin et al.,2005;Tang et al.,2008),尚无任何关于蝙蝠生态系统服务价值的量化研究(Voigt and Kingston,2016)。这种状况不仅不利于全面理解蝙蝠的生态作用,也十分不利于宣传蝙蝠的正面形象,尤其是在可能与蝙蝠相关的新冠肺炎等传染病流行时期。因此,我们认为中国蝙蝠研究者应该重视蝙蝠在害虫控制、种子传播和植物授粉方面的研究,量化相应的经济价值。并加强科普宣传,用事实和数据让政府和公众了解蝙蝠在生态系统和人类福祉中不可替代的作用。只有这样才能消除人们的误解,促使人们回归理性,并有助于中国蝙蝠物种多样性保护和开发“以蝠治虫”的绿色防控技术。
5.1.2 、加强分类学研究,厘清物种间关系
我国蝙蝠种类繁多、种群基数大,是维持生态平衡的重要类群(Wilson and Mittermeier,2019)。因此,加速推进蝙蝠分类学研究,明确中国蝙蝠物种多样性和本底分布状况具有重要意义。受限于国内蝙蝠分类学工作者数量较少,研究关注度不高,分类体系设置滞后,分类学疑难众多,故其资源与本底状况仍未明确,其中又以蝙蝠科中的森林型蝙蝠(如管鼻蝠亚科、伏翼属与类伏翼属、鼠耳蝠亚科等)研究最为不足。随着我国对生态环境保护的重视与生态环境的改善,以及全球气候变化、人类活动日益频繁及人兽共患重大疫情不断涌现的前提下,蝙蝠调查及分类学研究更具有必要性。通过传统形态学、几何形态学、系统发育基因组学、超声波特征、染色体分析等技术的综合运用,对物种的抢救性研究及对国内蝙蝠本底资源状况的充分掌握,不仅可完成对旧分类体系的修订,潜在性地发现新分布、新纪录种或新种,更可为日后的生态保护与修复、生物资源开发利用和重大疫情防控提供重要分类学支撑。同时,有机整合新的技术如机器学习乃至深度学习至分类学工作,加速分类成果的应用与转化,持续性资源投入与分类学人才培养,构建国内蝙蝠物种及标本资源信息网络交流平台,以及加强国内外更广泛的学术交流与合作都是十分必要。
5.1.3 、明确蝙蝠种群地位、动态和濒危等级
近年来,研究学者已经在中国很多地区开展了蝙蝠物种多样性调查,并已经基本掌握了其多样性和分布。然而,对于蝙蝠种群动态的长期监测,仅限于少数地区和少数常见或者具有一定特殊习性的物种。由于传统种群调查方法费时费力,加之蝙蝠对人为干扰极为敏感(Luo et al.,2012,2013),增加了蝙蝠种群调查的难度。随着音景生态学(Soundscape ecology)的发展,被动声学监测技术已经被广泛用来监测动物的多样性(Marques et al.,2012;Browning et al.,2017;Gibb et al.,2018),包括蝙蝠(Claireau et al.,2019)。2015年出台的北美蝙蝠监测计划(A Plan for the North American Bat Monitoring Program,NABat),主要利用声学方法来监测蝙蝠物种的地位和趋势(Loeb et al.,2015)。因此,我国研究者应广泛使用声学监测方法,结合传统的种群调查技术,长期监测蝙蝠多样性及其丰富度,进一步建立开放共享的蝙蝠物种多样性网络数据,才能为准确评估蝙蝠物种的濒危等级,并为蝙蝠物种多样性保护决策的制定提供科学依据。
5.1.4 、构建蝙蝠行为与保护之间的联系
近年来,动物行为学家开展了大量行为与物种保护关系的研究(Wedekind,2002),并推动形成了保护生物学的一个新的分支学科——保护行为学(Blumstein and Fernandez-Juricic,2004;李春旺等,2007)。对蝙蝠而言,声行为是其最重要的表型之一。因此,以声行为研究为切入点,揭示蝙蝠与环境间的相互作用,从而为蝙蝠的管理和保护提供理论指导。例如由于蝙蝠夜行性且飞行,野外种类鉴定和物种多样性监测都具有一定的难度,而蝙蝠回声定位声波具有种特异性,因此通过声学采样和机器学习等方法,根据回声定位声波进行种类鉴定与物种多样性监测,成为蝙蝠保护的有效手段(Mac Aodha et al.,2018)。另外,环境噪声可能导致蝙蝠回声定位、性选择和交流等行为的非适应性改变,进一步影响蝙蝠的繁殖成功和种群增长速率,因此对蝙蝠应对噪声干扰进行研究,能为蝙蝠物种多样性保护提供科学依据。
5.1.5 、保护基因组学与蝙蝠濒危物种保护
种群遗传多样性的丧失被认为是种群灭绝事件的先兆。随着基因组学的快速发展,保护基因组学已经被应用到野生动物的保护研究和实践。研究发现,通过比较1931年和2011—2012年采集的新加坡短耳犬蝠(Cynopterus brachyotis)41个个体的基因组序列,发现其种群遗传多样性大约在20世纪40年代持续下降,随后经历了一个明显的瓶颈效应,与人类世开始的时间一致(Chattopadhyay et al.,2019)。与1931年相比,短耳犬蝠的有效种群减少了近30倍,表明人类活动对蝙蝠种群的巨大影响,包括那些被认为对环境退化具有耐受性的物种(Chattopadhyay et al.,2019)。因此,开展蝙蝠保护基因组学研究非常必要,将为蝙蝠濒危物种的保护提供重要的指导和参考。
5.1.6 、加强蝙蝠病毒的研究
过去20年,我国在蝙蝠病毒的病原生态学研究已取得了长足进步。但我国幅员辽阔,地理景观与生物多样性丰富,随着研究的深入,我们发现当前已知的蝙蝠病毒仅是冰山一角。另外,虽然很多未知蝙蝠病毒逐渐浮出水面,但其潜在的公共卫生和生物学意义尚不清楚。因此,我们还需加大蝙蝠病毒的病原生态学、致病性,以及蝙蝠自身的免疫机制研究,预判病原的跨种传播,从而有效地维护公共卫生安全和保障人民健康。但在病毒生态研究过程中,存在滥杀蝙蝠取样的现象,建议相关部门尽快形成病毒溯源研究的行业规范,要求研究学者严谨科学地设计病毒溯源试验,最大程度上减少蝙蝠样本的采集数量。
5.1.7、 明确蝙蝠源人兽共患病传播的生态学机制
2019年末开始的新冠肺炎疫情再次将蝙蝠推向了科学研究和舆论的风口浪尖。“蝙蝠起源”假说是目前解释新型冠状病毒来源最主要的假说之一(Andersen et al.,2020;Zhou et al.,2020)。与SARS病毒的传播类似(Li et al.,2005;Hu et al.,2017),新型冠状病毒最可能的传播途径是:蝙蝠将冠状病毒传染给中间宿主,中间宿主再将变异后的病毒传播给人类。我国目前关于蝙蝠源冠状病毒跨物种传播机制的研究也主要集中在病毒多样性和病毒的受体结合能力两个方面,忽略了对病毒跨物种传播的生态学机制。
5.2 、保护蝙蝠栖息地
2000—2010年中国地表植被覆盖绿度逐渐提高,研究表明有约66%的区域呈增加趋势,表明整体上我国的绿度在增加(刘爽和宫鹏,2012)。植被保护对树栖蝙蝠非常重要,可为其提供足够可供选择的栖息地。与此同时,植被恢复之后,蝙蝠食物(果实、花蜜、昆虫等)也将进一步丰富,对蝙蝠种群存活和延续也起到关键作用。然而,洞穴旅游开发和城市现代化进程对蝙蝠栖息地丧失的影响仍是当前蝙蝠保护面临的主要问题。我们呼吁洞穴旅游开发必须进行严格的环评,特别关注对蝙蝠保护的影响。同时也呼吁相关部门重视古建筑物的保护,非必要的情况下尽量保留古建筑。
5.3 、建造人工蝙蝠栖息设施
随着快速城镇化,大量蝙蝠栖息地遭到了侵占或破坏(Feijó et al.,2019)。在此情况下,需要在人类居住的环境中主动为蝙蝠提供栖息场所,进而有效保护蝙蝠。首先,蝙蝠屋或蝙蝠箱由于建造成本相对低,安置灵活,占人类生存空间小等原因,往往成为理想的选择。蝙蝠屋或蝙蝠箱可建造在森林、农田等相对空旷的场所,在一些森林被大量砍伐或山洞被开发的地方,也建议安置不同大小的蝙蝠屋或蝙蝠箱。对于一些适宜蝙蝠潜在栖息的山洞或者废弃的矿洞,以及废弃的建筑等,可进行适当改造,作为蝙蝠的栖息场所。例如,可在洞口设置金属栅栏,既能让蝙蝠自由飞入飞出,又能有效防止无关人员进入。同时,在洞口设置相应的告示牌,给出警示以及相关联系人信息,在山洞管理单位授权和负责人监督下,规范相关的科学研究、洞内考察等活动。这些改造后的设施,可以作为蝙蝠临时或长久的栖息场所,从而有效地保护蝙蝠。
5.4、 加强蝙蝠保护公众教育
蝙蝠是夜行性动物,而且栖息地相对隐蔽,加之蝙蝠携带多种与人类新发传染病相关的病毒,人们便自然地把其与黑暗和邪恶联系在一起,迷信和缺乏了解导致人类大量杀害蝙蝠。除了专业人员的科学研究,提升公众对蝙蝠生态、经济和科研价值的认识,增强保护意识,促进普通民众、政府管理者和决策者参与蝙蝠保护活动,才能长期有效地改善蝙蝠保护的现状(Andrade and Rhodes,2012;Superina et al.,2018)。因此,建议依托中国野生动物保护协会等各级动物保护组织、中国动物学会等各级相关学术团体,以及各种民间动物保护组织和专业媒体等定期进行科普教育和宣传活动。此外,蝙蝠提供的生态服务价值也应更好地被量化和货币化,从而使公众对蝙蝠的经济价值有更理性的认知(Feijó et al.,2019)。
6 、结语
中国分布蝙蝠140余种,约占全球蝙蝠物种的10%,是世界上蝙蝠物种多样性最丰富的国家之一。虽然近50年以来,对中国蝙蝠物种的研究在多个学科领域已经取得了丰硕的成果,但直接与蝙蝠生态系统服务和保护相关的研究十分稀少。蝙蝠对环境变化极为敏感,全球和中国分别约22%和51%的蝙蝠物种位于近危等级之上,表明中国的蝙蝠物种受到更大程度的威胁。然而,除了少数物种之外,我们对中国大多数蝙蝠物种的种群地位和发展趋势仍不清楚。因此,我们应该鼓励学者、政府、企业、媒体和非政府组织等开展密切合作和知识共享,才能顺利实施各种保护措施,有效地促进中国蝙蝠物种多样性保护。
参考文献
Ahn M,Anderson D E,Zhang Q,Tan C W,Lim B L,Luko K,Wen M,Chia W N,Mani S,Wang L C,Ng J H J,Sobota R M,Dutertre C A,Ginhoux F,Shi Z L,Irving A T,Wang L F.2019.Dampened NLRP3-mediated inflammation in bats and implications for a special viral reservoir host.Nature Microbiology,4:789-799.
[]Aizpurua O,Budinski I,Georgiakakis P,Gopalakrishnan S,Ibaňez C,Mata V,Rebelo H,Russo D,Szodoray-Parádi F,Zhelyazkova V,Zrncic V,Gilbert M T P,Alberdi A.2018.Agriculture shapes the trophic niche of a bat preying on multiple pest arthropods across Europe:Evidence from DNA metabarcoding.Molecular Ecology,27(3):815-825.
Andersen K G,Rambaut A,Lipkin W I,Holmes E C,Garry R F.2020.The proximal origin of SARS-CoV-2.Nature Medicine,1-3.
Andrade G S M,Rhodes J R.2012.Protected areas and local communities:an inevitable partnership toward successful conservation strategies?Ecology and Society,17:14-23.
Andrews H.2018.Bat Roosts in Trees:A Guide to Identification and Assessment for Tree-Care and Ecology Professionals.Exeter:Pelagic Publishing.
Banerjee A,Baker M L,Kulcsar K,Misra V,Plowright R,Mossman K.2020.Novel insights into immune systems of bats.Frontiers in Immunology,11:26.
Bat Ecology,Evolution and Conservation Lab.2019.http://bat.nenu.edu.cn.cslg.naihes.cn/Collections/Samples.htm.
[]Bayat S,Geiser F,Kristiansen P,Wilson S C.2014.Organic contaminants in bats:trends and new issues.Environment International,63:40-52.
[]Bennett B S,Thies M L.2007.Organochlorine pesticide residues in guano of Brazilian free-tailed bats,Tadarida brasiliensis Saint-Hilaire,from East Texas.Bulletin of Environmental Contamination and Toxicology,78:191-194.
[]Blumstein D T,Fernandez-Juricic E.2004.The emergence of conservation behavior.Conservation Biology,18(5):1175-1177.
Boyles J G,Cryan P M,McCracken G F,Kunz T H.2011.Economic importance of bats in agriculture.Science,332(6025):41-42.
Brisbin I L.1966.Energy-utilization in a captive hoary bat.Journal of Mammalogy,47(4):719-720.
Browning E,Gibb R,Glover-Kapfer P,Jones K E.2017.Passive acoustic monitoring in ecology and conservation.Woking,United Kingdom:WWF.
Bu Y,Wang M,Zhang C,Zhang H,Niu H.2015.Study of roost selection and habits of a bat,Hipposideros armiger in mainland China.Pakistan Journal of Zoology,47(1):59-69.
[]Bumrungsri S,Harbit A,Benzie C,Carmouche K,Sridith K,Racey P.2008.The pollination ecology of two species of Parkia(Mimosaceae)in southern Thailand.Journal of Tropical Ecology,24(5):467-475.
[]Bumrungsri S,Sripaoraya E,Chongsiri T,Racey S P A.2009.The Pollination Ecology of Durian(Durio zibethinus,Bombacaceae)in Southern Thailand.Journal of Tropical Ecology,25(1):85-92.
Calisher C H.2015.Viruses in bats:a historic review.In:Wang L F,Cowled C eds.Bats and Viruses:A New Frontier of Emerging Infectious Diseases.Hoboken:John Wiley & Sons,Inc,23-45.
Chang Y,Song S J,Lin A Q,Zhang Y,Li Z L,Xiao Y H,Jiang T L,Feng J,Lin A Q.2019.The roles of morphological traits,resource variation and resource partitioning associated with the dietary niche expansion in the fish-eating bat Myotis pilosus.Molecular Ecology,28(11):2944-2954.
Chattopadhyay B,Garg K M,Mendenhall I H,Rheindt F E.2019.Historic DNA reveals Anthropocene threat to a tropical urban fruit bat.Current Biology,29(24):R1299-R1300.
Chen J,Liu T,Deng H,Xiao N,Zhou J.2017.A new species of Murina bats was discovered in Guizhou Province,China.Cave Research,2:1-10.
Chi T T,Liu M X,Tan X,Li Y,Xiao Y H,Sun K P,Jin L R,and Feng J.2019.Vocal development of Horsfield’s leaf-nosed bat pups(Hipposideros larvatus).Acta Chiropterologica,21:193-204.
Claireau F,Bas Y,Puechmaille S J,Julien J F,Allegrini B,Kerbiriou C.2019.Bat overpasses:an insufficient solution to restore habitat connectivity across roads.Journal of Applied Ecology,56(3):573-584.
[]Clarke F M,Pio D V,Race P A.2005.A comparison of logging systems and bat persity in the neotropics.Conservation Biology,19:1194-1204.
[]Cleveland C J,Betke M,Federico P,Frank J D,Hallam T G,Horn J.2006.Economic value of the pest control service provided bybrazilian free-tailed bats in south-central texas.Frontiers in Ecology & the Environment,4(5):238-243.
Craig P,Trail P,Morrell T E.1994.The decline of fruit bats in American Samoa due to hurricanes and overhunting.Biological Conservation,69:261-266.
[]Dong D,Lei M,Hua P,Pan Y H,Mu S,Zheng G,Pang E,Lin K,Zhang S.2016.The genomes of two bat species with long constant frequency echolocation calls.Molecular Biology and Evolution,34(1):20-34.
[]Eger J L,Lim B K.2011.Three new species of Murina from southern China(Chiroptera:Vespertilionidae).Acta Chiropterologica,13:227-243.
Feijó A,Wang Y Q,Sun J,Li F H,Wen Z X,Ge D Y,Xia L,Yang Q S.2019.Research trends on bats in China:a twenty-first century review.Mammalian Biology,98:163-172.
Feng L,Gao L,Lu H W,Muller R.2012.Noseleaf dynamics during pulse emission in horseshoe bats.PLoS ONE,7(5):e34685.
[]Feng Q,Li S,Wang Y X.2008.A new species of bamboo bat(Chiroptera:Vespertilionidae:Tylonycteris)from Southwestern China.Zoological Science,25(2):225-234.
[]Feng J,Chen M,Li Z X,Zhang S Y,Zhao H H,Zhou J.2002.Correlation between echolocation calls and morphological features among three kinds of bats.Progress in Natural Science,12:673-678.
Fenton M B.2001.Bats,Revised Edition.New York:Facts on File,Inc.
Fischer E M,Knutti R.2015.Anthropogenic contribution to global occurrence of heavy-precipitation and high-temperature extremes.Nature Climate Change,5:560-564.
Foley N M,Hughes G M,Huang Z,Clarke M,Jebb D,Whelan C V,Petit E J,Touzalin F,Farcy O,Jones G,Ransome R D,Kacprzyk J,O’Connell M J,Kerth G,Rebelo H,Rodrigues L,Puechmaille S J,Teeling E C.2018.Growing old,yet staying young:The role of telomeres in bats’ exceptional longevity.Science Advances,4:eaao0926.
Frick W F,Reynolds D S,Kunz T H.2010.Influence of climate and reproductive timing on demography of little brown myotis Myotis lucifugus.Journal of Animal Ecology,79:128-136.
Frick W F,Stepanian P M,Kelly J F,Howard K W,Kuster C M,Kunz T H,Chilson P B.2012.Climate and weather impact timing of emergence of bats.PLoS ONE,7:e42737.
Frick W F,Kingston T,Flanders J.2019.A review of the major threats and challenges to global bat conservation.Annals of the New York Academy of Sciences.
[]Fu Z Y,Bibikov N G,Wu F J,Chen Q C.2008.Duration selectivity of neurons in the inferior colliculus of the FM bat,Miniopterus magnate determined under different pulse repetition rate.Comparative Biochemistry and Physiology Part C:Toxicology & Pharmacology,148:454.
Fujita M S,Tuttle M D.1991.Flying foxes(chiroptera:pteropodidae):threatened animals of key ecological and economic importance.Conservation Biology,5(4):455-463.
Ge X Y,Li J L,Yang X L,Chmura A A,Zhu G J,Epstein J H,Mazet J K,Hu B,Zhang W,Peng C,Zhang Y J,Luo C M,Tan B,Wang N,Zhu Y,Crameri G,Zhang S Y,Wang L F,Daszak P,Shi Z L.2013.Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor.Nature,503(7477):535-538.
Gibb R,Browning E,Glover-Kapfer P,Jones K E.2018.Emerging opportunities and challenges for passive acoustics in ecological assessment and monitoring.Methods in Ecology and Evolution,10(2):169-185.
[]Guo W P,Lin X D,Wang W,Tian J H,Cong M L,Zhang H L,Wang M R,Zhou R H,Wang J B,Li M H,Xu J,Holmes E C,Zhang Y Z.2013.Phylogeny and origins of Hantaviruses harbored by bats,insectivores,and rodents.PLOS Pathogens,9:e1003159.
[]Guo W J,Yu W H ,Wang X Y,Csorba G,Li Y C,Wu Y.2017.First record of the collared sprite,Thainycteris aureocollaris (Chiroptera,Vespertilionidae)from China.Mammal Study,42(2):97-103.
Guo D G,Luo B,Zhang K K,Liu M X,Metzner W,Liu Y,Feng J.2019.Social vocalizations of big-footed myotis(Myotis macrodactylus)during foraging.Integrative Zoology,14:446-459.
Haddock J K,Threlfall C G,Law B,Hochuli D F.2019.Light pollution at the urban forest edge negatively impacts insectivorous bats.Biological Conservation,236:17-28.
Hage S R,Jiang T L,Berquist S W,Feng J,Metzner W.2013.Ambient noise induces independent shifts in call frequency and amplitude within the Lombard effect in echolocating bats.Proceedings of the National Academy of Sciences of the United States of America,110(10):4063-4068.
Hage S R,Jiang T L,Berquist S W,Feng J,Metzner W.2014.Ambient noise causes independent changes in distinct spectro-temporal features of echolocation calls in horseshoe bats.The Journal of Experimental Biology,217:2440-2444.
He F,Xiao N,Zhou J.2015a.A new species of Murina from China(Chiroptera:Vespertilionidae).Cave Research,2:2-6.
He B,Feng Y,Zhang H,Xu L,Yang W,Zhang Y,Li X,Tu C H.2015b.Filovirus RNA in fruit bats,China.Emerging Infectious Diseases,21:1675-1677.
He B,Zhang Y,Xu L,Yang W,Yang F,Feng Y,Xia L,Zhou J,Zhen W,Feng Y,Guo H,Zhang H,Tu C H.2014.Identification of perse alphacoronaviruses and genomic characterization of a novel severe acute respiratory syndrome-like coronavirus from bats in China.Journal of Virology,88:7070-7082.
Hedenstrom A,Johansson L C,Wolf M,von Busse R,Winter Y,Spedding G R.2007.Bat flight generates complex aerodynamic tracks.Science,316:894-897.
Heiker L M,Adams R A,Ramos C V.2018.Mercury bioaccumulation in two species of insectivorous bats from urban china:Influence of species,age,and land use type.Archives of Environmental Contamination and Toxicology,75:585-593.
Heim O,Lorenz L,Kramer-Schadt S,Jung K,Voigt C C,Eccard J A.2017.Landscape and scale-dependent spatial niches of bats foraging above intensively used arable fields.Ecological Processes,6(1):24.
[]Hoffmann M,Belant J L,Chanson J S,Cox N A,Stuart S N.2011.The changing fates of the world’s mammals.Philosophical Transactions of the Royal Society B,366(1578):2598-2610.
Hoffmann M,Hilton-Taylor C,Angulo A,et al.2010.The impact of conservation on the status of the world’s vertebrates.Science,330:1503-1509.
[]Hong W,Zhao H.2014.Vampire bats exhibit evolutionary reduction of bitter taste receptor genes common to other bats.Proceedings of the Royal Society B:Biological Sciences,281(1788):20141079.
[]Hu B,Zeng L P,Yang X L,Ge X Y,Zhang W,Li B,Xie J Z,Shen X R,Zhang Y Z,Wang N,Luo D S,Zheng X S,Wang M N,Daszak P,Wang L F,Cui J,Shi Z L.2017.Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus.PLOS Pathogens,13:e1006698.
Hu Y,Cheng H,Tao S.2016.The challenges and solutions for cadmium-contaminated rice in China:a critical review.Environment International,92-93:515-532.
Huang C J,Yu W H,Xu Z X,Qiu Y X,Chen M,Qiu B,Motokawa M,Harada M,Li Y C,Wu Y.2014.A cryptic species Ty-lonycteris fulvidus within Tylonycteris pachypus group(Chiroptera:Vespertilionidae)and its population genetic structure in southern China and nearby regions.International Journal of Biological Sciences,10(2):200-211.
International Union for the Conservation of Nature(IUCN).2020.The IUCN Red List of Threatened Species.Version 2020-1.http://h-s.www.iucnredlist.org.Downloaded on 19 March 2020.
Jakobsen L,Olsen M N,Surlykke A.2015.Dynamics of the echolocation beam during prey pursuit in aerial hawking bats.Proceedings of the National Academy of Sciences,112:8118-8123.
[]Jiang T L,Feng J,Sun K P,Wang J.2008.Coexistence of two sympatric and morphologically similar bat species Rhinolophus affinis and Rhinolophus pearsoni.Progress in Natural Science,18(5):523-532.
[]Jiang T L,Lu G J,Sun K P,Luo J H,Feng J.2013.Coexistence of Rhinolophus affinis and Rhinolophus pearsoni revisited.Acta Theriologica,58(1):47-53.
Jiang T L,Huang X B,Wu H,Lin H J,Müller R,Hoyt J R,Feng J.2016.Autumn flight activity of the greater horseshoe bat at hibernacula.Animal Biology,66(2):119-131.
Jiang T L,Guo X,Lin A,Wu H,Sun C,Feng J,Kanwal J S.2019a.Bats increase vocal amplitude and decrease vocal complexity to mitigate noise interference during social communication.Animal Cognition,22(2):199-212.
Jiang T L,Wang J,Wu H,Csorba G,Puechmaille S J,Benda P,Boireau J,Toffoli R,Courtois J Y,Nyssen P,Colombo P,Feng J.2019b.The patterns and possible causes of global geographical variation in the body size of the greater horseshoe bat(Rhinolophus ferrumequinum).Journal of Biogeography,46:2363-2377.
[]Jiang T,Wu H,Feng J.2015.Patterns and causes of geographic variation in bat echolocation pulses.Integrative Zoology,10:241-256.
[]Jiao H,Wang Y,Zhang L,Jiang P,Zhao H.2018.Lineage-specific duplication and adaptive evolution of bitter taste receptor genes in bats.Molecular Ecology,27(22):4475-4488.
Jiao H,Zhang L,Xie H W,Simmons N B,Liu H,Zhao H.2019.Trehalase gene as a molecular signature of dietary persification in mammals.Molecular Biology and Evolution,36(10):2171-2183.
[]Jin L R,Feng J,Sun K P,Liu Y,Wu L,Li Z X,Zhang X C.2005.Foraging strategies in the greater horseshoe bat(Rhinolophus ferrumequinum)on lepidoptera in summer.Chinese Science Bulletin,50(14):1477-1482.
[]Jin L R,Yang S L,Kimball R T,Xie L F,Yue X K,Luo B,Sun K P,Feng J.2015.Do pups recognize maternal calls inpomona leaf-nosed bats,Hipposideros pomona?Animal Behaviour,100:200-207.
Jones K E,Patel N G,Levy M A,Storeygard A,Balk D,Gittleman J L,Daszak P.2008.Global trends in emerging infectious diseases.Nature,451:990-993.
Jones G,Jacobs D S,Kunz T H,Willig M R,Racey P A.2009.Carpe noctem:the importance of bats as bioindicators.Endangered Species Research,8:93-115.
[]Kannan K,Yun S H,Rudd R J,Behr M.2010.High concentrations of persistent organic pollutants including PCBs,DDT,PBDEs and PFOS in little brown bats with white-nose syndrome in New York,USA.Chemosphere,80:613-618.
[]Karp D S,Daily G C.2014.Cascading effects of insectivorous birds and bats in tropical coffee plantations.Ecology,95(4):1065-1074.
Koh J,Itahana Y,Mendenhall I H,Low D,Soh E X Y,Guo A K,Chionh Y T,Wang L F,Itahana K.2019.ABCB1 protects bat cells from DNA damage induced by genotoxic compounds.Nature Communications,10:2820.
Kunz T H,Stern A A.1995.Maternal investment and post-natal growth in bats.Symposia of Zoological Society of London,67:123-138.
[]Kunz T H,Whitaker J O,Wadanoli M D.1995.Dietary energetics of the insectivorous Mexican free-tailed bat(Tadarida brasiliensis)during pregnancy and lactation.Oecologia(Berlin),101(4):407-415.
Kunz T H,Torrez E B D,Bauer D,Lobova T,Fleming T H.2011.Ecosystem services provided by bats.Annals of the New York Academy of Sciences,1223(1):1-38.
[]Kuo H C,Fang Y P,Csorba G,Lee L L.2006.The definition of Harpiola(Vespertilionidae:Murininae)and the description of a new species from Taiwan.Acta Chiropterologica,8:11-19.
[]Kuo H C,Fang Y P,Csorba G,Lee L L.2009.Three New Species of Murina(Chiroptera:Vespertilionidae)from Taiwan.Journal of Mammalogy,90:980-991.
[]Kuo H C,Soisook P,Csorba G,Wang C N,Rossiter S J.2017.A taxonomic revision of the Kerivoula hardwickii complex(Chiroptera:Vespertilionidae)with the description of a new species.Acta Chiropterologica,19(1):19-39.
Lagunas-Rangel F A.2020.Why do bats live so long?Possible molecular mechanisms.Biogerontology,21:1-11.
Lau S K P,Woo P C Y,Li K S M,Huang Y,Tsoi H W,Wong B H L,Wong S S Y.Leung S Y,Chan K H,Yuen K Y.2005.Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats.Proceedings of the National Academy of Sciences of the United States of America,102:14040-14045.
LaVal R K.2004.Impact of global warming and locally changing climate on tropical cloud forest bats.Journal of Mammalogy,85:237-244.
Li G,Jones G,Rossiter S J,Chen S F,Parsons S,Zhang S Y.2006.Phylogenetics of small horseshoe bats from East Asia based on mitochondrial DNA sequence variation.Journal of Mammalogy,87(6):1234-1240.
Li G,Wang J,Rossiter S J,Jones G,Cotton J A,Zhang S.2008.The hearing gene Prestin reunites echolocating bats.Proceedings of the National Academy of Sciences of the United States of America,105(37):13959-13964.
Li G,Wang J,Rossiter S J,Jones G,Zhang S.2007.Accelerated FoxP2 evolution in echolocating bats.PLoS ONE,2(9):e900.
Li L,Chi H,Liu H,Xia Y,Irwin D M,Zhang S,Liu Y.2018.Retention and losses of ultraviolet-sensitive visual pigments in bats.Scientific Reports,8(1):11933.
Li W,Shi Z,Yu M,Ren W,Smith C,Epstein J H,Wang H,Crameri G,Hu Z,Zhang H,Zhang J,McEachern J,Field H,Daszak P,Eaton B T,Zhang S,Wang L F.2005.Bats are natural reservoirs of SARS-like coronaviruses.Science,310:676-679.
[]Li Y,Liu Z,Shi P,Zhang J.2010.The hearing gene Prestin unites echolocating bats and whales.Current Biology,20(2):R55-R56.
[]Li Y Y,Liu Z,Qi F Y,Zhou X,Shi P.2016.Functional effects of a retained ancestral polymorphism in Prestin.Molecular Biology and Evolution,34(1):88-92.
[]Lin H J,Kanwal J S,Jiang T L,Liu Y,Feng J.2015.Social and vocal behavior in adult greater tube-nosed bats(Murina leucogaster).Zoology,118:192-202.
Liu H Q,Wei J K,Li B,Wang M S,Wu R Q,Rizak J D,Zhong L,Wang L,Xu F Q,Shen Y Y,et al.2015.Divergence of dim-light vision among bats(order:Chiroptera)as estimated by molecular and electrophysiological methods.Scientific Reports,5:11531.
[]Liu S,Yu W,Li F,Zhao J,Yin R,Zhou Z.2018.Fertilizer application in rural cropland drives cadmium enrichment in bats dwelling in an urban area.Environmental Pollution,242:970-975.
[]Liu Y Y,Wang Y M,Zhang Z X,Bu Y Z,Niu H X.2018.Roost selection and ecology of stoliczka’s trident bat,Aselliscus stoliczkanus(hipposideridae,chiroptera)in China.Mammalian Biology Zeitschrift Für S?ugetierkunde,95:143-149.
[]Liu Y,Cotton J A,Shen B,Han X,Rossiter S J,Zhang S.2010.Convergent sequence evolution between echolocating bats and dolphins.Current Biology,20(2):R53-R54.
Liu Y,Han N,Franchini L F,Xu H,Pisciottano F,Elgoyhen A B,Rajan K E,Zhang S.2012a.The voltage-gated potassium channel subfamily KQT member 4(KCNQ4)displays parallel evolution in echolocating bats.Molecular Biology and Evolution,29(5):1441-1450.
Liu Y,He G,Xu H,Han X,Jones G,Rossiter S J,Zhang S.2014.Adaptive functional persification of lysozyme in insectivorous bats.Molecular Biology and Evolution,31(11):2829-2835.
Liu Y,Xu H,Yuan X,Rossiter S J,Zhang S.2012b.Multiple adaptive losses of alanine-glyoxylate aminotransferase mitochondrial targeting in fruit-eating bats.Molecular Biology and Evolution,29(6):1507-1511.
Liu Y,Zhang S,Zhao J,Zhang F,Hu R.2013.Isolation of Irkut virus from a Murina leucogaster bat in China.PLoS Neglected Tropical Diseases,7:e2097.
Liu Z,Li S,Wang W,Xu D,Murphy R W,Shi P.2011.Parallel evolution of KCNQ4 in echolocating bats.PLoS ONE,6(10):e26618.
Liu Z,Qi F Y,Zhou X,Ren H Q,Shi P.2014.Parallel sites implicate functional convergence of the hearing gene Prestin among echolocating mammals.Molecular Biology and Evolution,31(9):2415-2424.
Loeb S C,Rodhouse T J,Ellison L E,Lausen C L,Richard J D,Irvine K M,Ingersoll T E,Coleman J T H,Thogmartin W E,Sauer J R,Francis C M,Bayless M L,Stanley T R,Johnson D H.2015.A plan for the North American Bat monitoring program(NABat).General Technical Report SRS-208.Asheville,NC,USA:United States Department of Agriculture,Forest Service,Southern ResearchStation [www document].URL www.srs.fs.usda.gov/pubs/gtr/gtr_srs208.pdf.
[]Lundy M,Montgomery I,Russ J.2010.Climate change-linked range expansion of Nathusius’ pipistrelle bat,Pipistrellus nathusii(Keyserling & Blasius,1839).Journal of Biogeography,37:2232-2242.
Luo B,Leiser-Miller L,Santana S E,Zhang L,Liu T,Xiao Y H,Liu Y,Feng J.2019.Echolocation call pergence in bats:a comparative analysis.Behavioral Ecology and Sociobiology,73(11):154.
[]Luo B,Lu G J,Chen K,Guo D G,Huang X B,Liu Y,Feng J.2017.Social calls honestly signal female competitive ability in Asian particoloured bats.Animal Behaviour,127:101-108.
Luo B,Santana S E,Pang Y L,Wang M,Xiao Y H,Feng J.2019.Wing morphology predicts geographic range size in vespertilionid bats.Scientific Reports,9:4526.
Luo F,Metzner W,Wu F J,Zhang S Y,Chen Q C.2008.Duration-sensitive neurons in the inferior colliculus of horseshoe bats:Adaptations for using cf-fm echolocation pulses.Journal of Neurophysiology,99:284-296.
[]Luo J H,Lu G J,Feng J.2012.Diurnal capture reduces the colony size of Hipposideros armiger(Chiroptera:Hipposideridae).Ma-mmalia,76:447-449.
[]Luo J H,Jiang T L,Lu G J,Wang L,Wang J,Feng J.2013.Bat conservation in China:should protection of subterranean habitats be a priority?Oryx,47:526-531.
Luo J H,Koselj K,Zseb?k S,Siemers B M,Goerlitz H R.2014.Global warming alters sound transmission:differential impact on the prey detection ability of echolocating bats.Journal of the Royal Society Interface,11:20130961.
Luo J H,Goerlitz H R,Brumm H,Wiegrebe L.2015a.Linking the sender to the receiver:vocal adjustments by bats to maintain signal detection in noise.Scientific Reports,5(1):18556.
[]Luo J H,Siemers B M,Koselj K.2015b.How anthropogenic noise affects foraging.Global Change Biology,21(9):3278-3289.
[]Luo J H,Hage S R,Moss C F.2018.The Lombard effect:from acoustics to neural mechanisms.Trends in Neuroscience,41:938-949.
[]Ma J,Jones G,Zhang S Y,Shen J X,Metzner W,Zhang L B.2010.Dietary analysis confirms that rickett’s big-footed bat(Myotis ricketti)is a piscivore.Journal of Zoology,261(3):245-248.
Ma J,Zhang J S ,Zhang L B,Zhang S Y,Metzner W.2006a.Dietary characteristics of Myotis ricketti in Beijing,north China.Journal of Mammalogy,87(2):339-344.
[]Ma J,Kobayasi K,Zhang S Y,Metzner W.2006b.Vocal communication in adult greater horseshoe bats,Rhinolophus ferrumequinum.Journal of Comparative Physiology A,192:535-550.
Mac Aodha O,Gibb R,Barlow K E,Browning E,Firman M,Freeman R,Harder B,Kinsey L,Mead G R,Newson S E,Pandourski I,Parsons S,Russ J,Szodoray-Paradi A,Szodoray-Paradi F,Tilova E,Girolami M,Brostow G,Jones K E.2018.Bat detective-Deep learning tools for bat acoustic signal detection.PLoS Computational Biology,14(3):e1005995.
[]Maine J J,Boyles J G.2015.Bats initiate vital agroecological interactions in corn.Proceedings of the National Academy of Sciences,112(40):12438-12443 .
Marais D J D,Mitchell J M,Meinschein W G,Hayes J M.1980.The carbon isotope biogeochemistry of the inpidual hydrocarbons in bat guano and the ecology of the insectivorous bats in the region of carlsbad,new mexico.Geochimica Et Cosmochimica Acta,44(12):2075-2086.
[]Marques T A,Thomas L,Martin S W,Mellinger,D K,Ward J A,Moretti D J,Harris D,Tyack P L.2012.Estimating animal po-pulation density using passive acoustics.Biological Reviews,88(2):287-309.
McConkey K R,Drake D R,Franklin J,Tonga F.2004.Effects of Cyclone Waka on flying foxes(Pteropus tonganus)in the Vava’u Islands of Tonga.Journal of Tropical Ecology,20:555-561.
McConkey K R,Drake D R.2015.Low redundancy in seed dispersal within an island frugivore community.AoB Plants,7:plv088.
McCracken G F,Bernard R F,Gamba-Rios M,Wolfe R,Krauel J J,Jones D N,Russell A L,Brown V A.2018.Rapid range expansion of the Brazilian free-tailed bat in the southeastern United States,2008-2016.Journal of Mammalogy,99:312-320.
McCracken G F,Gillam E H,Westbrook J K,Lee Y F,Jensen M L,Balsley B B.2008.Brazilian free-tailed bats(Tadarida brasiliensis:Molossidae,Chiroptera)at high altitude:links to migratory insect populations.Integrative and Comparative Biology,48(1):107-118.
Moratelli R,Calisher C H.2015.Bats and zoonotic viruses:can we confidently link bats with emerging deadly viruses?Mem Inst Oswaldo Cruz,110:1-22.
[]Muscarella R,Fleming T H.2007.The role of frugivorous bats in tropical forest succession.Biological Reviews,82(4):573-590.
[]Niu H,Wang N,Zhao L,Liu J.2007.Distribution and underground habitats of cave-dwelling bats in China.Animal Conservation,10(4):470-477.
[]Park K J.2015.Mitigating the impacts of agriculture on biopersity:bats and their potential role as bioindicators.Mammalian Biology,80(3):191-204.
Peng K,Peng Y J,Wang J,Yang M J,Fu Z Y,Tang J,Chen Q C.2017.Latency modulation of collicular neurons induced by electric stimulation of the auditory cortex in Hipposideros pratti:In vivo intracellular recording.PLoS ONE,12:e0184097.
[]Pierson E D,Elmqvist T,Rainey W E,Cox P A.1996.Effects of tropical cyclonic storms on flying fox populations on the south Pacific Islands of Samoa.Conservation Biology,10(2):438-451.
Pimm S L,Jenkins C N,Abell R,Brooks T M,Gittleman J L,Joppa L N,Raven P H,Roberts C M,Sexton J O.2014.The biopersity of species and their rates of extinction,distribution,and protection.Science,344(6187):1246752.
Roberts B J,Catterall C P,Eby P,Kanowski J.2012.Long-distance and frequent movements of the flying-fox Pteropus poliocephalus:implications for management.PLoS ONE,7:e42532.
[]Sachanowicz K,Wower A,Bashta A T.2006.Further range extension of Pipistrellus kuhlii(Kuhl,1817)in central and eastern Europe.Acta Chiropterologica,8:543-548.
[]Scanlon A T,Petit S,Tuiwawa M,Naikatini A.2018.Response of primary and secondary rainforest flowers and fruits to a cyclone,and implications for plant-servicing bats.Global Change Biology,24:3820-3836.
Seluanov A,Gladyshev V N,Vijg J,Gorbunova V.2018.Mechanisms of cancer resistance in long-lived mammals.Nature Reviews Cancer,18:433-441.
[]Shen Y Y,Liang L,Li G S,Murphy R W,Zhang Y P.2012.Parallel evolution of auditory genes for echolocation in bats and toothed whales.PLoS Genetics,8(6):e1002788.
Shen Y Y,Liu J,Irwin D M,Zhang Y P.2010.Parallel and convergent evolution of the dim-light vision gene RH1 in bats(Order:Chiroptera).PLoS ONE,5(1):e8838.
[]Sherwin H A,Montgomery W I,Lundy M G.2012.The impact and implications of climate change for bats.Mammal Review,43:171-182.
Shi B Y,Wang Y Z,Gong L X ,Chang Y,Liu T,Zhao X,et al.2020.Correlation of skull morphology and bite force in a bird-eating bat(Ia io:Vespertilionidae).Frontiers in Zoology,17:8.
Song S J,Lin A Q,Jiang T L,Zhao X,Metzner W,Feng J.2019.Bats adjust temporal parameters of echolocation pulses but not those of communication calls in response to traffic noise.Integrative Zoo-logy,14:576-588.
[]Stahlschmidt P,Brühl C A.2012.Bats at risk?Bat activity and insecticide residue analysis of food items in an apple orchard.Environmental Toxicology and Chemistry,31(7):1556-1563.
[]Stepanian P M,Wainwright C E.2018.Ongoing changes in migration phenology and winter residency at Bracken Bat Cave.Global Change Biology,24:3266-3275.
[]Stone E L,Harris S,Jones G.2015.Impacts of artificial lighting on bats:a review of challenges and solutions.Mammalian Biology,80(3):213-219.
Sun C N,Zhang C M,Gu H,Jiang T L,Feng J.2019.Self-assessment strategy during contest decisions between male Great Himalayan leaf-nosed bats.Behavioral Ecology and Sociobiology,73(4).DOI:10.1007/s00265-019-2657-0
[]Sun K P,Feng J,Jiang T L,Ma J,Zhang Z Z,Jin L R.2008.A new cryptic species of Rhinolophus macrotis(Chiroptera:Rhinolophidae)from Jiangxi Province,China.Acta Chiropterologica,10(1):1-10.
Sun X,Jen P H S.1986.Ear position affects the auditory space representation in the inferior colliculus of bats.The Journal of the Acoustical Society of America,80:S107-S108.
Sun X,Jen P H S.1989.Electrical stimulation of bat superior colliculus influences responses of inferior collicular neurons to acoustic stimuli.Brain Research,497:214-222.
Superina M,Cortés Duarte,A,Trujillo F.2018.Connecting research,management,education and policy for the conservation of armadillos in the Orinoco Llanos of Colombia.Oryx,53:17-26.
[]Tang J,Wei C X,Chen M X,Wang Q C,Kong H F,Fu Z Y,Chen Q C.2017.Recovery cycle of inferior collicular neurons in Hipposideros pratti under behavior-related sound stimulus and the best Doppler-shift compensation conditions.Physiology & Behavior,171:236-242.
Tang Z H,Cao M,Sheng L X,Ma X F,Zhang S Y.2008.Seed dispersal of morus macroura(moraceae)by two frugivorous bats in Xishuangbanna,S W China.Biotropica,40(1):127-131.
Teeling E C,Springer M S,Madsen O,Bates P,O’Brien S J,Murphy W J.2005.A molecular phylogeny for bats illuminates biogeo-graphy and the fossil record.Science,307:580-584.
[]Tiunov M P,Kruskop S V and Feng J.2011.A New Mouse-Eared Bat(Mammalia:Chiroptera,Vespertilionidae)from South China.Acta Chiropterologica,13(2):271-278.
Voigt C C,Kingston T.2016.Bats in the Anthropocene:Conservation of Bats in a Changing World.Springer International Publishing.
Wang H,Zhao H,Sun K,Huang X,Jin L,Feng J.2019.Evolutionary basis of high-frequency hearing in the cochleae of echolocators revealed by comparative genomics.Genome Biology and Evolution,12(1):3740-3753.
Wang J,Gao W,Wang L,Metzner W,Ma J,Feng J.2010.Seasonal variation in prey abundance influences habitat use by greater horseshoe bats(Rhinolophus ferrumequinum)in a temperate deci-duous forest.Canadian Journal of Zoology,88:315-323.
[]Wang J,Kanwal J,Zhang C L,Jiang T L,Lu G J,Feng J.2010.Seasonal habitat use by greater horseshoe bat Rhinolophus ferrumequinum(Chiroptera:Rhinolophidae)in Changbai Mountain temperate forest,northeast China.Mammalia,74(3):257-266.
[]Wang Q,Qi J,Yuan Y,Xuan Y,Han P,Wan Y,Ji W,Li Y,Wu Y,Wang J,Iwamoto A,Woo Patrick C Y,Yuen K Y,Yan J,Lu G,Gao George F.2014.Bat origins of MERS-CoV supported by bat coronavirus HKU4 usage of human rceptor CD26.Cell Host & Microbe,16:328-337.
[]Wang X Y,Guo W J,Yu W H,Csorba G,Motokawa M,Li F,Zhang Q P,Zhang C Y,Li Y C,Wu Y.2017.First record and phylogenetic position of Myotis indochinensis (Chiroptera,Vespertilionidae)from China.Mammalia,81(6):605-609.
[]Wanger T C,Darras K,Bumrungsri S,Tscharntke T,Klein A M.2014.Bat pest control contributes to food security in Thailand.Biological Conservation,171(1):220-223.
[]Wedekind C.2002.Sexual selection and life-history decisions:Implications for supportive breeding and the management of captive populations.Conservation Biology,16:1204-1211.
[]Welbergen J A,Klose S M,Markus N,Eby P.2008.Climate change and the effects of temperature extremes on Australian flying-foxes.Proceedings of the Royal Society:Biological Sciences,275:419-425.
Wilson D E,Mittermeier R A.2019.Handbook of the Mammals of the World.vol.9.Bats.Barcelona:Lynx Edicions.
Wohlgemuth M J,Luo J,Moss C F.2016.Three-dimensional auditory localization in the echolocating bat.Current Opinion in Neurobiology,41:78-86.
Wu F,Zhao S,Yu B,Chen Y M,Wang W,Song Z G,Hu Y,Tao Z W,Tian J H,Pei Y Y,Yuan M L,Zhang Y L,Dai F H,Liu Y,Wang Q M,Zheng J J,Xu L,Holmes E C,Zhang Y Z.2020.A new coronavirus associated with human respiratory disease in China.Nature,579:265-269.
Wu H,Jiang T L,Liu S,Lu G J,Feng J.2020.Acoustic identification of two morphologically similar bat species,Miniopterus magnater and Miniopterus fuliginosus(Chiroptera,Miniopteridae).Mammalia,84(2):201-206.
[]Wu J,Jiao H,Simmons N B,Lu Q,Zhao H.2018.Testing the sensory trade-off hypothesis in New World bats.Proceedings of the Ro-yal Society B:Biological Sciences,285(1885):20181523.
[]Wu Y,Harada M,Morokawa M.2009.Taxonomy of Rhinolophus yunanensis Dobson,1872(Chiroptera:Rhinolophidae)with a description of a new species from Thailand.Acta Chiropterologica,11(2):237-246.
[]Wu Y,Li Y C,Lin L K,Harada M,Chen Z,Motokawa M.2012.New Records of Kerivoula titania(Chiroptera:Vespertilionidae)from Hainan Island and Taiwan.Mammal Study,37:69-72.
[]Wu Y,Motokawa M,Harada M,Thong V D,Lin L K,Li Y C.2012.Morphometric Variation in the pusillus Group of the Genus Rhinolophus(Mammalia:Chiroptera:Rhinolophidae)in East Asia.Zoological Science,29(6):396-402.
[]Wu Y,Motokawa M,Harada M.2008.A new species of horseshoe bat of the genus Rhinolophus from China(Chiroptera:Rhinolophidae).Zoological Science,25(4):438-443.
[]Wu Y,Thong V D.2011.A New Species of Rhinolophus(Chiroptera:Rhinolophidae)from China.Zoological Science,28(3):235-241.
[]Xiao Y H,Wang L,Hoyt J R,Jiang T L,Lin A Q,Feng J.2018.Stereotypy and variability of social calls among clustering female big-footed myotis(Myotis macrodactylus).Zoological Research,39:114-122.
[]Xie J,Li Y,Shen X,Goh G,Zhu Y,Cui J,Wang L F,Shi Z L,Zhou P.2018.Dampened STING-dependent interferon activation in bats.Cell Host Microbe,23(3):297-301.
Xu L,Wu J,Li Q,Wei Y,Tan Z,Cai J,Guo H,Yang L E,Huang X,Chen J,Zhang F,He B,Tu C H.2019.Seroprevalence,cross antigenicity and circulation sphere of bat-borne hantaviruses revealed by serological and antigenic analyses.PLoS Pathogens,15:e1007545.
Yang M J,Peng K,Wang J,Tang J,Fu Z Y,Wang X,Chen Q C.2018.Amplitude and duration sensitivity of single-on and double-on neurons to CF-FM stimuli in inferior colliculus of Pratt’s roundleaf bat(Hipposideros pratti).Journal of Comparative Physiology A,204:653-665.
Yang X L,Tan C W,Anderson D E,Jiang R D,Li B,Zhang W,Zhu Y,Lim X F,Zhou P,Liu X L,Guan W,Zhang L,Li S Y,Zhang Y Z,Wang L F,Shi Z L.2019.Characterization of a filovirus(Měnglà virus)from Rousettus bats in China.Nature Microbiology,4:390-395.
Yang Y,Du L,Liu C,Wang L,Ma C,Tang J,Baric R S,Jiang S,Li F.2014.Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirus.Proceedings of the National Academy of Sciences of the United States of America,111:12516.
Yu W H,Li F,Csorba G,Xu,Z X,Wang X Y,Guo W J,Li Y C,Wu Y.2018.A revision of Kerivoula hardwickii and occurrence of K.furva(Chiroptera:Vespertilionidae)in China.Zootaxa,4461:45-56.
[]Yu W H,Csorba G,Wu Y.2020.Tube-nosed variations-a new species of the genus Murina(Chiroptera:Vespertilionidae)from China.Zoological Research,41(1):70-77.
Zeng X,Chen J,Deng H,Xiao N,Zhou J.2018.A new species of Murina from China(Chiroptera:Vespertilionidae).Ekoloji,103:9-16.
[]Zhang J,Sun X,Jen P H S.2001.Corticofugal modulation of frequency tuning of inferior collicular neurons in big brown bat,Eptesicus fuscus.China Science Bulletin,46:836-839.
Zhang L B,Jones G,Rossiter S,Ades G,Liang B,Zhang S Y.2005.Diet of flat-headed bats,Tylonycteris pachypus and T.robustula,in Guangxi,south China.Journal of Mammalogy,86(1):61-66.
Zhang J S,Han N J,Jones G,Lin L K,Zhang J P,Zhu G J,Huang D W,Zhang S Y.2007.A new species of Barbastella(Chiroptera:Vespertilionidae)from North China.Journal of Mammalogy,88(6):1393-1403.
[]Zhang L B,Jones G,Zhang J S,Zhu G J,Parsons S,Rossiter S J,Zhang S Y.2009.Recent surveys of bats(Mammalia:Chiroptera)from China.Ⅰ.Rhinolophidae and Hipposideridae.Acta Chiropterologica,11(1):71-88.
[]Zhang L,Zhu G,Jones G,Zhang S.2009.Conservation of bats in China:problems and recommendations.Oryx,43(2):179-182.
Zhang J P,Jen P H S,Sun X.2000.Direction-dependent corticofugal modulation of frequency-tuning curves of inferior collicular neurons in the big brown bat,Eptesicus fuscus.Journal of Comparative Physiology A,186:913-922.
[]Zhang J S,Gareth J,Zhang L B,Zhu G J,Zhang S Y.2010.Recent surveys of bats(Mammalia:Chiroptera)from China Ⅱ.Pteropodidae.Acta Chiropterologica,12(1):103-116.
[]Zhang L,Sun K P,Liu T,Zhao H B,Csorba G,Jin L R,Thong V D,Feng J.2018.Multilocus phylogeny and species delimitation within the philippinensis group(Chiroptera:Rhinolophidae).Zoologica Scripta,47:655-672.
[]Zhang C M,Jiang T L,Lu G J,Lin A Q,Sun K P,Liu S,Feng J.2018.Geographical variation in the echolocation calls of bent-winged bats,Miniopterus fuliginosus.Zoology,131:36-44.
Zhang K K,Liu T,Liu M X,Li A Q,Xiao Y H,Metzner W,Liu Y.2019a.Comparing context-dependent call sequences employing machine learning methods:an indication of syntactic structure of greater horseshoe bats.Journal of Experimental Biology,222:24.
Zhang Y,Lin A Q,Ding J N,Yang X Y,Jiang T L,Liu Y,Feng J.2019b.Performance of Doppler shift compensation in bats varies with species rather than with environmental clutter.Animal Beha-viour,158:109-120.
Zhao H,Rossiter S J,Teeling E C,Li C,Cotton J A,Zhang S.2009.The evolution of color vision in nocturnal mammals.Proceedings of the National Academy of Sciences of the United States of America,106(22):8980-8985.
Zhao H,Zhou Y,Pinto C M,Charles-Dominique P,Galindo-Gonzalez J,Zhang S,Zhang J.2010.Evolution of the sweet taste receptor gene Tas1r2 in bats.Molecular Biology and Evolution,27(11):2642-2650.
Zhao H,Xu D,Zhang S,Zhang J.2012.Genomic and genetic evidence for the loss of umami taste in bats.Genome Biology and Evolution,4(1):73-79.
Zhao H B.2020.COVID-19 drives new threat to bats in China.Science,367(6485):1436.
Zhao X,Jiang T L,Liu H,Wang Y Z,Liu Y,Feng J.2019.Acoustic signalling of aggressive intent in the agonistic encounters of female Asian particoloured bats.Animal Behaviour,149:65-75.
Zhou P,Fan H,Lan T,Yang X L,Shi W F,Zhang W,Zhu Y,Zhang Y W,Xie Q M,Mani S,Zheng X S,Li B,Li J M,Guo H,Pei G Q,An X P,Chen J W,Zhou L,Mai K J,Wu Z X,Li D,Anderson D E,Zhang L B,Li S Y,Mi Z Q,He T T,Cong F,Guo P J,Huang R,Luo Y,Liu X L,Chen J,Huang Y,Sun Q,Zhang X L L,Wang Y Y,Xing S Z,Chen Y S,Sun Y,Li J,Daszak P,Wang L F,Shi Z L,Tong Y G,Ma J Y.2018.Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin.Nature,556:255-258.
Zhou P,Yang X L,Wang X G,Hu B,Zhang L,Zhang W,Si H R,Zhu Y,Li B,Huang C L,Chen H D,Chen J,Luo Y,Guo H,Jiang R D,Liu M Q,Chen Y,Shen X R,Wang X,Zheng X S,Zhao K,Chen Q J,Deng F,Liu L L,Yan B,Zhan F X,Wang Y Y,Xiao G F,Shi Z L.2020.A pneumonia outbreak associated with a new coronavirus of probable bat origin.Nature,579:270-273.
[]Zhou Y,Dong D,Zhang S,Zhao H.2009.Positive selection drives the evolution of bat bitter taste receptor genes.Biochemical Gene-tics,47(3-4):207-215.
[]Zhou Z M,Guillén S A,Lim B K,Eger J L,Wang Y X,Jiang X L.2009.A new species from southwestern China in the Afro-Palearctic lineage of the horseshoe bats(Rhinolophus).Journal of Mammalogy,90(1):57-73.
[]Smith A T,解焱.2009.中国兽类野外手册.长沙:湖南教育出版社.
[]王应祥.2003.中国哺乳动物种和亚种分类名与分布大全.北京:中国林业出版社.
[]王宗禕,汪松.1962.青海发现的大狐蝠(Pteropus giganteus Brünnich).动物学报,14(4):494.
[]王静,王新华,江廷磊,王磊,卢冠军,由玉岩,刘颖,李丹,冯江.2010.马铁菊头蝠捕食活动与猎物资源的关系.兽类学报,30(2):157-162.
[]邓亚东,陈伟海,张远海,陈旺,易旭敏.2011.岩溶洞穴旅游开发对生态环境影响分析——以巴马水晶宫为例.桂林理工大学学报,31(3):412-417.
[]由玉岩.2013.栖息洞穴干扰对特有种大卫鼠耳蝠种群数量和基因丰富度的影响.生物学杂志,30(2):28-32.
[]冯庆,王应祥,林苏.2007.中国安氏长舌果蝠的分类记述.动物学研究,28(6):647-653.
[]冯庆,蒋学龙,李松,王应祥.2006.中国翼手类一属、种新纪录.动物分类学报,31(1):224-230.
[]冯江,李振新,张喜臣.2001.我国蝙蝠保护研究现状及对策.东北师大学报(自然科学版),33(2):70-75.
[]刘元.2015.20世纪60年代党对林业政策的调整.湘潭大学硕士学位论文.
[]刘少英,吴毅.2019.中国兽类图鉴.福州:海峡书局.
[]刘伟,王延校,何新焕,牛红星.2011.太行山南段洞栖蝙蝠的分布及栖息地重要性分析.兽类学报,31(4):371-379.
[]刘志霄,张佑祥,张劲硕,张礼标.2014.湖南省发现艾氏管鼻蝠.动物学杂志,49:132-135.
[]刘爽,宫鹏.2012.2000-2010年中国地表植被绿度变化.科学通报,16:53-64.
[]江廷磊,刘颖,冯江.2008.中国翼手类一新纪录种.动物分类学报,33(1):212-216.
[]孙心德,Jen P H S,张季平,章森福.1992.菊头幅出生后下丘听神经元反应特性的演化.生物物理学报,8:683-690.
[]孙淙南,郭熊,黄晓宾,江廷磊,冯江.2016.环境噪声对蝙蝠回声定位叫声及飞行活动的影响.兽类学报,36:397-403.
[]寿振黄.1958.东北兽类调查报告.北京:科学出版社.
[]寿振黄.1962.中国经济动物志:兽类.北京:科学出版社.
[]李春旺,蒋志刚,张恩权,古远.2007.保护行为学:正在兴起的保护生物学分支学科.生物多样性,15(3):312-318.
[]吴毅,杨奇森,夏霖,彭洪元,周昭敏.2004.中国蝙蝠新记录——马氏菊头蝠.动物学杂志,39(5):109-110.
[]吴毅,陈子禧,王晓云,黎舫,胡宜峰,郭伟健,余文华,李玉春.2017.哈氏管鼻蝠在广东的新发现及南岭树栖蝙蝠物种多样性.广州大学学报(自然科学版),16:1-7.
[]吴毅,郑福军,李艳,陈莹,肖玲.2004a.广州地区濒危物种扁颅蝠Tylonycteris pachypus的种群数量变化与环境因素的关系.中山大学学报(自然科学版),43(5):91-94.
[]吴毅,原田正史,李艳红.2004b.四川七种蝙蝠的核型.兽类学报,24(1):30-35.
[]何晓瑞,杨白仑.1991.中国翼手类一新记录——泰国狐蝠.兽类学报,11(1):71-72.
汪冬梅.2003.中国城市化问题研究.北京:中国经济出版社.
[]张礼标,张劲硕,梁冰,张树义.2004.中国翼手类新记录——小巨足蝠.动物学研究,25(6):556-559.
[]张礼标,梁冰,周善义,卢立仁,张树义.2004.扁颅蝠与褐扁颅蝠的集群结构.动物学报,50(3):326-333.
[]张礼标,巩艳艳,朱光剑,洪体玉,赵旭东,毛秀光.2010.中国翼手目新记录——马来假吸血蝠.动物学研究,31(3):328-332.
[]张劲硕,张礼标,赵辉华,梁冰,张树义.2005b.中国翼手类新记录——小褐菊头蝠.动物学杂志,40(2):96-98.
张劲硕.2010.中国蝙蝠(哺乳纲:翼手目)分类的综合研究.博士学位论文,中国科学院研究生院,148-152.
[]张树义.2009.我国近年来关于蝙蝠的研究进展及对未来的展望.中国科学基金,23(5):265-268.
[]罗一宁.1987.我国兽类新记录——缺齿鼠耳蝠.兽类学报,7(2):159-159.
[]罗峰,梁冰,张礼标,张树义.2005.食虫蝙蝠与昆虫之间的相互作用和协同进化关系.动物学杂志,40(3):104-110.
[]金龙如,冯江,孙克萍,刘颖,吴磊,李振新,张喜臣.2005.马铁菊头蝠夏季对鳞翅目昆虫的捕食策略.科学通报,50(13):1334-1339.
[]周晓明,孙心德.1997.早期声音暴露对蝙蝠中脑下丘听神经元频率调谐特性发育的影响.动物学研究,18:60-65.
[]周晓明,孙心德.1999.GABA及bicuculline对幼年鼠耳蝠下丘神经元听空间反应特性的影响.中国神经科学杂志,15:268-273.
[]赵辉华,张树义,周江,刘自民.2002.中国翼手类新记录——高鞍菊头蝠.兽类学报,22(1):74-76.
[]草林娣.2007.论中国园林的蝙蝠符号——福寿德善美仁的象征.苏州教育学院学报,24:5-9.
[]胡宜峰,余文华,岳阳,黄正澜懿,李玉春,吴毅.2019.海南岛翼手目物种多样性现状与分布预测.生物多样性,27(4):400-408.
[]夏武平.1964.中国动物图谱:兽类.北京:科学出版社.
[]党飞红,余文华,王晓云,郭伟健,庄卓升,梅廷媛,张秋萍,李锋,李玉春,吴毅.2017.中国渡濑氏鼠耳蝠种名订正.四川动物,36(1):7-13.
[]钱燕文,张洁,汪松,关贯赉,沈孝宙.1965.新疆南部的鸟兽.北京:科学出版社.
[]徐丽静,孙心德.1998.γ-氨基丁酸、谷氨酸等对蝙蝠中脑下丘薄片神经元诱发电活动的影响.生物物理学报,14:93-97.
[]郭熊,赵健伦,江廷磊,冯江.2015.东方蝙蝠在交通噪声环境中的回声定位行为.动物学杂志,50(4):615-620.
[]龚小燕,黄太福,吴涛,张佑祥,彭清忠,刘志霄.2018.西南鼠耳蝠(Myotis altarium)栖息生态学.生态学报,38(21):7549-7556.
蒋志刚,马勇,吴毅,王应祥,周开亚.2015.中国哺乳动物多样性和地理分布.北京:科学出版社.
[]蒋志刚,江建平,王跃招,张鹗,张雁云,李立立,谢锋,蔡波,曹亮,郑光美等.2016.中国脊椎动物红色名录,生物多样性,24(5):500-551.
[]简耘,石磊,李丹,张纯淳,石苗苗,王嵘,陈小勇.2008.森林砍伐对苦槠种群遗传结构的影响.生态学报,28(11):1-7.
[]谭敏,朱光剑,洪体玉,叶建平,张礼标.2009.中国翼手类新记录——小蹄蝠.动物学研究,30(2):204-208.
[]熊郁良.1975.昆明花红洞地区几种蝙蝠的生态观察.动物学报,21(4):48-55.
[]潘清华,王应祥,岩崑.2007.中国哺乳动物彩色图鉴.北京:中国林业出版社.





