摘 要: 为了探讨长江江豚(Neophocaena asiaeorientalis asiaeorientalis)种群的生存动态和管护对策,本研究利用漩涡模型(Vortex 10.3.7.0)对长江江豚种群生存力进行了回顾和现状分析。结果发现,以1999年作为回顾分析起点,长江江豚的灭绝概率最可能为0.0754,且不高于0.5629。平均灭绝时间最可能为93.5年,且不少于87.9年。以2017年作为现状分析起点,长江江豚种群灭绝概率为0.3551~0.6985,平均灭绝时间为83.5~88.0年。在特定情景下,设定新生幼豚雄性所占比例分别为0.55、0.50和0.45,经模拟计算得出,雄性化让灭绝概率由0.6985增加到0.9157,雌性化使其下降到0.3664。同时,数据比较发现,新生幼豚雄性所占比例下降10%,成年雌豚生殖率提高10%,二者最终效果相当。上述研究结果标明,长江江豚种群生存风险较大,重点保护,修复其生境,尤其是母子豚相关生境有利于长江江豚的保护。
关键词: 长江江豚; 种群生存力; 旋涡模型; 死亡率; 生殖率;
Abstract: To study the population dynamics and conservation strategies of the Yangtze finless porpoise (Neophocaena asiaeorientalis asiaeorientalis), Vortex (v.10.3.7.0) was used to analyze the population viability. Taking 1999 as the starting point, a retrospective analysis revealed that the extinction probability of finless porpoise was most likely to be 0.0754 and was not higher than 0.5629. The mean extinction time was most likely to be 93.5 years and not less than 87.9 years. Taking 2017 as the starting point, through the analysis of the current situation, it was found that the extinction probability of the Yangtze finless porpoise population was 0.3551–0.6985, and the average extinction time was 83.5–88.0 years. In certain situations, the proportion of newborn male Yangtze finless porpoise was 0.55, 0.50, and 0.45, respectively. The results showed that masculinization increased the probability of extinction from 0.6985 to 0.9157, and feminization brought it down to 0.3664. At the same time, comparison of the data found that a 10% decrease in the proportion of newborn males and a 10% increase in the breeding rate of adult females had the same effect. The above results suggested that the extinction risk of the finless porpoise population in the Yangtze River was relatively high. Focusing on the protection and restoration of high-quality habitats, especially those related to parents–children, is important for the protection of the Yangtze finless porpoise.
Keyword: Yangtze finless porpoise; PVA; Vortex model; Mortality; Breeding;
种群生存力分析(Population Viability Analysis, PVA)是一种以物种生活史信息为参数,运用计算机模型来估计濒危物种种群未来大小和灭绝风险的一种方法,多用于模拟孤立小种群的生存动态过程,可以预测种群数量变化和灭绝概率(Brook et al, 2000a、b)。应用这一理论,发展了许多计算机应用程序,包括RAMAS(Ferson et al, 1989)、NEMESIS(Gilpin, 1993)、ALEX(Possingham et al, 1992)和Vprtex (Lacy et al, 1993)等,其中,Vortex模型(漩涡模型)应用最为广泛。国内学者已经利用该模型对白鳍豚(Lipotes vexillifer) (张先锋等, 1994)、长江江豚(Neophocaena asiaeorientalisasiaeorientalis) (张先锋等, 1999)、普氏野马(Equus przewalskii) (裴鹏祖等, 2018)、川金丝猴(Rhinopithecus roxellana) (张宇等, 2018)、猕猴(Macaca mulatta) (楚原梦冉等, 2019)等种群动态进行模拟和研究。张先锋等(1999)采用漩涡模型(Vortex 7.3)模拟长江江豚的种群生存力,预测了长江江豚未来的种群动态。20年来,长江江豚种群结构发生了新变化,种群生态学研究也取得了新成果(杨健等, 2000; 于道平等, 2005; 张新桥, 2011; Kimura et al, 2012; Mei et al, 2014、2017; Zhou et al, 2018; Yuan et al, 2018),有必要对长江江豚种群生存状况进行再评估和再认识。为此,本研究利用漩涡模型(Vortex 10.3.7.0)对长江江豚种群生存力及其致危因素进行分析,以期为未来种群管理和有效保护提供有益参考。

1、 种群生存力分析的种群参数及其估计
1.1 、种群描述
长时间以来,长江和海洋江豚的分类关系一直存在争议,曾被认为是同一个物种(3个亚种),后被认定为2个物种:以台湾海峡为界,以南海域是印太江豚;以北海域包括长江流域为窄脊江豚(Neophocaena asiaeorientalis)。2种江豚形态上区分的显着特点是其脊背宽窄和疣粒数量多少。长江江豚是江豚的最濒危的一个亚种,仅生活在长江中下游干流和大型通江湖泊—鄱阳湖和洞庭湖中。但目前基于江豚种群基因组学最新研究结果显示,极度濒危的长江江豚种群应被作为一个独立的物种(Zhou et al, 2018)。长江江豚不仅是全世界唯一一种生存于淡水环境的鼠海豚科(Phocaenidae)动物,也是长江流域极可能唯一现存的的水生哺乳动物(2007年白鱀豚功能性灭绝)。自上世纪九十年代以来,长江江豚的数量呈现出持续性的衰减趋势,2018年7月正式发布《2017长江江豚生态科学考察报告》,这也是目前最新的长江江豚科考成果。江豚种群数量大幅下降的趋势得到遏制,估算长江江豚数量约为1012头,其中,鄱阳湖约为457头,是野外江豚分布密度最高的区域。长江江豚发生了一些新变化,种群趋于向受人为扰动较少的江段集中,过半数水域种群持续下降或没有发现种群,种群分布呈碎片化特征(先义杰, 2018)。因其受威胁的严峻程度,长江江豚已被按照国家一级保护动物进行保护。2018年,中国科学院深海科学与工程研究所海洋哺乳动物研究团队对长江江豚进行了全基因组测序,组装得到了2.22 Gb的基因组草图,对江豚基因组的研究奠定了基础(Yuan et al, 2018)。
本研究以1999年作为回顾分析的起点,2017年作为模拟验证时间节点和现状分析的起点,进行长江江豚种群生存力分析。为进一步提升精度和可信度,将迭代次数设定为10000,为前人同类研究的10倍。MVP定义为以95%的概率健康存活100年所需的最小种群数量。估算方法为:分析不同初始种群数量(Initial population size, Ni)在100年内的种群生存力,当种群存活概率达到95%时的Ni即为MVP(Lacy et al, 2009、2017)。
1.2 、迁移扩散
长江江豚与海江豚之间存在着显着而稳定的遗传分化,提示它们之间缺乏基因交流而出现了生殖隔离,长江江豚因此被认定为独立物种,长江江豚和海江豚最明显的差别是一个生活在淡水,另一个生活在咸水。通过大数据分析,发现长江江豚和海江豚在进化上显着不同,特别是基因上体现对水环境的适应性差异,表明了长江江豚和其他江豚之间,已经不可能出现基因交流和种群间混合,因此,不存在迁移扩散问题(Zhou et al, 2018)。
1.3 、种群繁殖参数
长江江豚为混交制,雌性可能先于雄性性成熟,雌性首次生育年龄为4~6龄,雄性为4.5~7龄(郝玉江等, 2006)。Shirakihara等(1993)发现,2头21龄和23龄的个体可以怀孕和哺乳。长江江豚无论野外还是人工环境下均无产双胞胎的记录,因此,在模拟中设定每胎最大产仔数为1。长江江豚新生婴豚的性比为1∶1,雌性首次生育年龄为4龄,雄性为5龄,最高繁殖年龄约为20龄。该物种的种群增长应为密度制约型,即参加繁殖的雌性数占全部成年雌性数的比例[P(N)]随种群大小(N)的变化而变化。Fowler(1981)认为,描述这种变化强度的参数B取值2能够更好地模拟密度制约型种群的增长,描述雌性交配率降低的参数A取值也为2。假设N接近环境容纳量K时,繁殖雌性的比例P(K)为25%;而当N接近0时,P(0)为70%(张先锋等, 1999)。
1.4 、繁殖率
张先锋等(1992)研究指出,长江江豚繁殖周期一般为2年,年生殖率为20%。2020年1月公开报道显示,湖北石首天鹅洲迁地保护区内,江豚种群5年内净增长108%。因此,推测自然环境下,成年雌豚生殖率可能为20%~25%。目前,关于长江江豚流产率和幼豚夭折率以及流产或幼豚夭折等对其繁殖周期的影响还不全面,进一步深入研究有助于提升相关参数估算的精度。
1.5、 死亡率
张先锋等(1999)研究发现,长江江豚0~1和1~2两个年龄组,死亡率为20%,其他年龄组的死亡率为15%。梅志刚等(2011)研究指出,20世纪90年代以来,其种群下降速率每年约为6.3%,并推测幼体死亡率较高可能是决定其种群动态的重要因素。在模拟中设定0~1年龄组,死亡率为20%~30%;1~2年龄组,死亡率为20%;其他年龄组为15%。
1.6 、灾害
张先锋等(1999)设置2种灾害类型,发生的频率为10%,以死亡率均值为存活率下降率(95%),成年雌性死亡率表示繁殖率下降率(95%)。2016年1月,***总书记在重庆召开推动长江经济带发展座谈会,提出要把修复长江生态环境摆在压倒性位置,“共抓大保护、不搞大开发”。中国农业农村部发布《关于长江流域重点水域禁捕范围和时间的通告》,通告中指出,长江干流和重要支流除水生生物自然保护区和水产种质资源保护区以外的天然水域,最迟自2021年1月1日0时起实行暂定为期10年的常年禁捕,期间禁止天然渔业资源的生产性捕捞。人类的不利于长江江豚保护的活动会减少,但像2008年的大雪灾害、2019年的鄱阳湖大干旱极低水位以及意外事故等灾害仍然存在。我们综合病害等生理风险和意外等生态风险2种灾害类型,同时长江江豚种群存在碎片化分布趋势,影响程度应该更大。为此,以1999年作为回顾分析的起点,2017年为模拟验证时间节点,设置灾害2种类型,发生的频率为10%,以死亡率均值为存活率下降率(95%),成年雌性死亡率表示繁殖率下降率(95%)。以2017年作为现状分析的起点,设置灾害2种类型,发生的频率均为10%,以死亡率均值为存活率下降率(90%),成年雌性死亡率表示繁殖率下降率(90%)。
1.7 、初始头数及环境容纳量
张先锋等(1999)设置长江江豚的初始头数为2000头,环境容纳量为5000头,每5年环境容纳量将减少10%。为此,以1999年作为回顾分析的起点,2017年为模拟验证时间节点,也设置初始头数为2000头,环境容纳量为5000头。环境容纳量每年减少2%~3%,连续减少15年。以2017年作为现状分析的起点,设置初始头数为1012头,环境容纳量为4000头,每年增加1%,连续变动10年。
1.8 、近亲繁殖
对于小种群而言,近交衰退是影响其长期存活的重要因素之一。Vortex软件通过模拟有无近亲繁殖时的种群动态来说明其对种群的影响。Ralls等(1988)研究了40个哺乳动物种群致死等价系数,认为每个二倍体有3.14个致死基因当量。因此,设置3.14作为致死等价系数。
1.9、 人为捕获和补充
目前,没有对长江江豚的捕获和补充,故该参数设置为0。
2、 结果与分析
2.1、 不同模拟情景下长江江豚种群生存力回顾分析
通过Vortex模拟计算可知,以1999年作为回顾分析的起点,8种不同情景下具体结果见表1。长江江豚种群未来100年的内禀增长率r为–0.0622~ –0.0231,周限增长率λ为0.9397~0.9701,净生殖率Ro为0.5747~0.8211,平均世代时间雌性为8.55~8.90年,雄性为9.41~9.73年,平均现存种群数量为8.20±6.08~184.16±139.75头,平均种群数量为2.03±4.36~183.92±139.81头,遗传多样性为0.6990± 0.1455~0.9585±0.0301。灭绝概率为0.0013~0.7828,平均首次灭绝时间为84.4~95.4年,具体见表2。以2017年长江江豚数量约为1012头作为模拟验证来看,情景2和情景4与实际情况不吻合,为小概率情景。情景3模拟结果与实际调查情况契合度最好,情景1次之,情景5也较理想。情景5和情景7种群存活概率达到95%以上。
表1 长江江豚种群生存力回顾分析的不同模拟情景
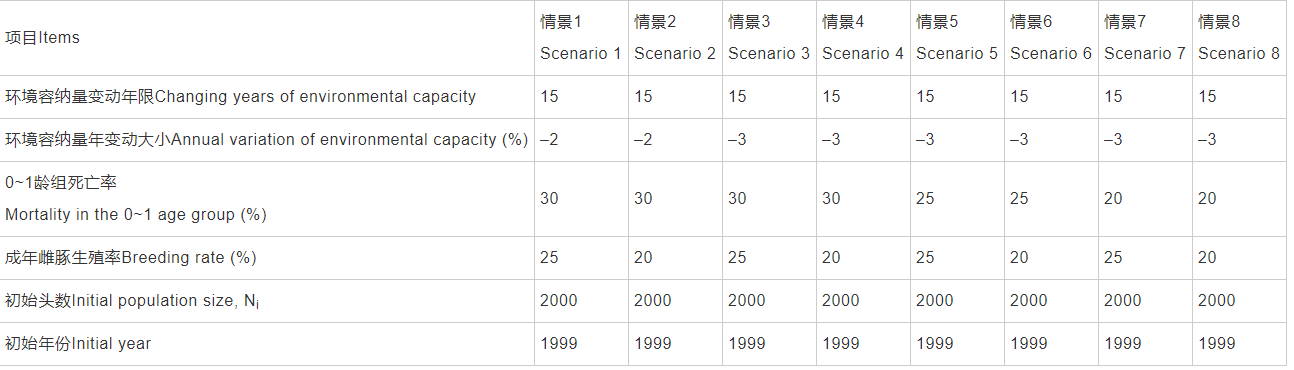
表2 长江江豚种群生存力回顾分析结果
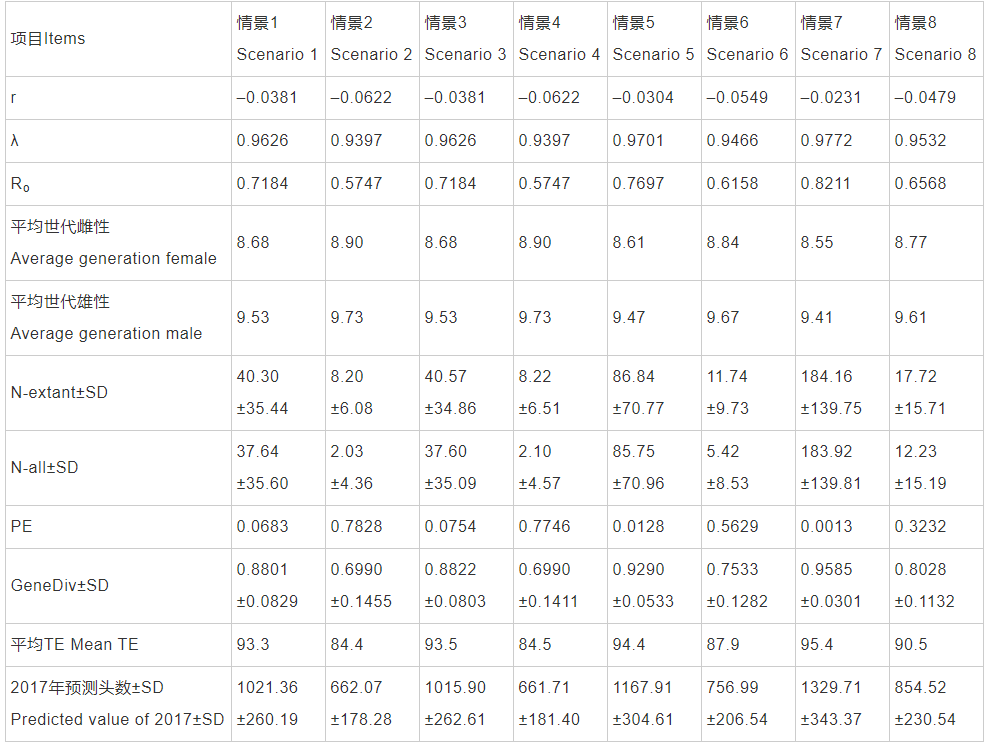
注:r: 内禀增长率; λ: 周限增长率; Ro: 净生殖率; N-extant: 平均现存种群数量; PE: 平均种群数量; GeneDiv: 灭绝概率; TE: 遗传多样性和首次灭绝时间(年)。表5同Note: r: Annual internal growth rate; λ: Finite rate of increase; Ro: Net reproduction rate; N-extant: Average number of existing populations; PE: Average population number; GeneDiv: Extinction probability; TE: Genetic diversity and first extinction time (years). Same in the Tab.5
2.2、 长江江豚种群最小可存活种群估计
情景5的模拟结果与实际调查情况契合度较理想,种群灭绝概率为0.0128。因此,情景5可以作为阶段性的科学研究及精准管护的目标。为此,求取情景5条件下的最小可存活种群(Minimum viable population, MVP),初始头数为1000开始模拟,按照需要每次增减5头,不难发现,情景5时,MVP为1305头,具体结果见表3。同理,采取渐进逼近的方法发现,情景5条件下,满足MVP为1305头的最小环境容量为2300头。
2.3 、不同模拟情景下长江江豚种群生存力现状分析
首先,在回顾分析中情景3基础上,以2017年作为现状分析的起点,设置初始头数为1012头,环境容纳量为5000头和4000头,发现灭绝概率分别为0.7012和0.6950。在情景5的基础上,设定环境容纳量为4000头,环境容纳量分别每年变动–1%、0%和1%,连续变动10年,发现灭绝概率分别为0.4635、0.4592和0.4588。随后,环境容纳量设定为4000头,每年增加1%,连续变动10年,以2017年作为起点进行不同模拟情景的生存动态分析,具体见表4。长江江豚种群100年的内禀增长率r为–0.0733~ –0.0207,周限增长率λ为0.9293~0.9795,净增值率Ro为0.5235~0.8413,平均世代雌性为8.36~8.83年,雄性为9.23~9.66年,平均现存种群数量5.49±3.58~ 110.81±108.10头,平均种群数量为0.11±0.80~ 107.91±108.10头,遗传多样性为0.5801±0.1671~ 0.9169±0.0659。灭绝概率为0.0264~0.9860,平均首次灭绝时间为68.4~91.9年,具体见表5。
表3 特定情景下长江江豚种群MVP模拟分析
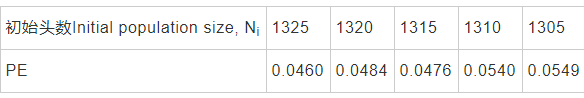
表4 长江江豚种群生存力现状分析的不同模拟情景

表5 长江江豚种群生存力现状分析结果

在现状分析情景6的基础,设定性比雄性化、雌性化偏离10%,即雄性所占比例分别为0.55、0.50和0.45,模拟计算得出,长江江豚种群100年的内禀增长率r分别为–0.0608、–0.0493和–0.0386,周限增长率λ分别为0.9410、0.9519和0.9621,净增值率Ro分别为0.5889、0.6543和0.7198,平均世代雌性分别为8.71、8.61和8.51年,雄性分别为9.56、9.46和9.37年,平均现存种群数量分别为7.30±5.69、11.67± 10.17和22.42±22.58头,平均种群数量分别为0.76± 2.66、3.75±7.66和14.42±20.39头,遗传多样性分别为0.6490±0.1597、0.7191±0.1483和0.7950±0.1237。灭绝概率为0.9157、0.6985和0.3664,平均首次灭绝时间为76.5、83.5和88.0年。
3 、讨论
本研究首先以1999年作为回顾分析的起点,2017年为模拟验证时间节点,在综合分析运用历史文献资料以及最新研究成果的基础上,围绕环境容纳量、0~1龄组死亡率、成年雌豚生殖率设定了回顾分析8种不同情景,长江江豚种群100年灭绝概率为0.0013~0.7828,平均灭绝时间为84.4~95.4年。以2017年长江江豚数量约为1012头作为模拟验证来看,情景2和情景4与实际情况不吻合,情景3模拟结果与实际调查情况吻合度最好,情景1次之,情景5也较理想。情景5和情景7的种群存活概率达到95%以上。提示上述情景3中模拟参数与历史生境及长江江豚种群特性的契合度较好,在该情景下进行深入系统分析具有较强的理论和实际价值。通过Vortex模拟计算可知,环境容纳量为5000头和4000头,发现灭绝概率分别为0.7012和0.6950。进一步设定环境容纳量为4000头,环境容纳量分别每年变动–1%、0%和1%,连续变动10年,发现灭绝概率分别为0.4635、0.4592和0.4588,提示环境容纳量小范围变动对长江江豚种群生存力影响不大,结果差异不大。从回顾分析结果来看,长江江豚的灭绝概率最可能为0.0754,且不高于0.5629。平均灭绝时间最可能为93.5年,且不少于87.9年。0~1龄组死亡率最可能为30%左右,成年雌豚生殖率最可能为25%左右。杨光等(1998)研究指出,长江江豚0~2龄组死亡率为0.308,繁殖率可高达0.7。张先锋等(1999)研究发现,长江江豚0~1龄组的死亡率为20%,年生殖率为20%。因此,模拟计算确认的长江江豚0~1龄组死亡率最可能为30%左右,成年雌豚生殖率最可能为25%左右,在合理范围之内。通过进一步分析,不难发现,成年雌豚生殖率和0~1龄组死亡率与种群生存力关系密切,模拟计算出,成年雌豚生殖率为25%时,长江江豚0~1龄组死亡率由30%分别下降为25%和20%时,灭绝概率由0.0754下降为0.0128和0.0013。0~1龄组死亡率为30%时,成年雌豚生殖率由25%下降为20%时,灭绝概率由0.0754增高为0.7746。因此,长江江豚种群所面临灭绝“漩涡”,直接原因应该是“一低一高”,即成年雌豚生殖率低,而0~1龄组死亡率高,但除了长江江豚自身生理生态以及进化演化适应方面的原因,长江江豚的生境干扰、破坏,特别是适合母子豚栖息、抚幼以及幼豚生长发育生境的数量减少和质量下降,导致其种群被动碎片化分布,个体水平可能有利于减小生存压力。但从种群水平来说,繁殖、遗传方面等风险加剧,“一低一高”现象持续恶化。另一方面,饵料鱼类资源下降,特别是渔汛等减少,捕食成本大幅提高,而长江江豚作为淡水水生哺乳动物,能量需求相对较大,特别是求偶、怀孕和哺乳等特定生理阶段,饵料相对不足,必将严重影响繁殖活动,并对幼仔生存带来巨大挑战,最终可能让长江江豚陷入种群灭绝“漩涡”。在回顾分析中,求取了情景5,即最可能达到情景下的MVP,为1305头,且满足该条件的最小环境容纳量为2300头。提示在进行长江江豚保护时,长江干流、鄱阳湖以及洞庭湖等水域在加强重点水域重点保护的同时,必须强化长江中下游水域大范围的“互联互通”,单一特定水域难以承载长江江豚的持续生存繁衍。
本研究中,从0~1龄组死亡率和成年雌豚生殖率的角度设定了8种不同模拟情景,环境容纳量设定为4000头,每年增加1%,连续变动10年,以2017年作为现状分析的起点,从上述回顾分析的结果来看,现状分析中的情景6和情景7是最可能情景,即长江江豚种群灭绝概率可能为0.3551~0.6985,平均灭绝时间可能为83.5~88.0年。而情景1和情景5是小概率情景,情景2和情景3是可能性较低情景,情景4和情景8是相对理想情景。长江江豚新生幼豚性比(雄性所占比例)对模拟结果具有较大影响,性比雄性化、雌性化偏离10%,灭绝概率由0.6985分别增加为0.9157和下降为0.3664。提示长江江豚新生幼豚性比研究对于长江江豚保护具有特别意义,通过比较相关数据,现状分析中情景6新生幼豚性比雌性化10%,与情景7模拟结果相当,即在情景6时,新生幼豚性比雌性化10%与成年雌豚生殖率提高10%,效果是相当的,提示人为诱导甚至控制长江江豚新生幼豚性比可能是进行长江江豚抢救性保护的措施之一。当种群面临灭绝漩涡风险时,重点保护、修复母子豚栖息区等特定自然生境,缓解甚至逆转长江江豚成年雌豚生殖率低,而0~1龄组死亡率高的现状,有利于长江江豚的保护,当然这需要对长江江豚生理生态学、行为生态学、繁殖生态学等进行深入而系统的研究。需要特别指出的是,我国实施伏季休渔制度已有20多年,在缓解捕捞对渔业资源带来的巨大压力和资源养护方面发挥了积极作用(胡芷君等, 2019),长江十年禁捕等战略性政策的落地,必将有效增加长江江豚饵料资源,对其保护意义重大。本文模型使用的参数是由系统整理前人长江江豚研究结果获得的,可能存在一定的误差,导致预测准确性降低,精度还有进一步提高的空间,但本文首次运用迭代1万次的方法进行相关模拟和估计,迭代次数为前人研究的10倍,一定程度上减小了系统误差,能够为长江江豚种群生存力分析及相关濒危机制探讨,特别是长江江豚种群的保护提供一定借鉴和管护依据。同时,有报道使用灰色预测模型,分析影响资源丰度变化因子,并建立多种资源丰度预报模型,以此反映该种类的资源丰度动态,应用效果较好,提示多种模型协同应用可能是未来种群生存力分析的发展方向之一(闫永斌等, 2019)。
(编辑 冯小花)
参考文献
[1] Brook BW, Burgman M, Frankham R. Differences and congruencies between PVA packages: The importance of sex ratio for predictions of extinction risk. Conservation Ecology, 2000a, 4(1): 920
[2] Brook BW, O’Grady JJ, Chapman AP, et al. Predictive accuracy of population viability analysis in conservation biology. Nature, 2000b, 404(6776): 385–387
[3] Chuyuan MR, Zan QJ, Yang Q, et al. Population dynamic and viability analysis of Rhesus macaque (Macaca mulatta) in Neilingding Nature Reserve, Guangdong Province. Chinese Journal of Wildlife, 2019, 2: 259–266 [楚原梦冉, 昝启杰, 杨琼, 等. 广东内伶仃岛猕猴种群动态及种群生存力分析. 野生动物学报, 2019, 2: 259–266]
[4] Ferson S, Ginzburg L, Silvers A. Extreme event risk analysis for age-structured populations. Ecological Modelling, 1989, 47(1–2): 175–187
[5] Fowler CW. Density dependence as related to life history strategy. Ecology, 1981, 62: 602–610
[6] Gaskin DE, Smith GJD, Watson AP, et al. Reproduction in the porpoise Phocoeridae: Implication for manayement. Repint Whalcann, 1984(S6): 135–148
[7] Gilpin M. NEMESIS. Version 1.0 users guide. Conservation biology simulations: Process simulations of dynamics at the level of genes, individuals, populations and metapopulations. Michael Gilpin, 1993, 419 W. Harrison, Bozeman, Montana 59715 USA
[8] Hu ZJ, Shan XJ, Yang T. et al. Preliminary evaluation of summer fishing moratorium in the Bohai Sea. Progress in Fishery Sciences [胡芷君, 单秀娟, 杨涛, 等. 渤海伏季休渔效果初步评价. 渔业科学进展. https://doi.org/10.19663/ j.issn2095-9869.20190623001]
[9] Hao YJ, Wang D, Zhang XF. Review on breeding biology of Yangtze finless porpoise (Neophocaena phocaenoides asiaeorientalis). Acta Theriologica Sinica of China, 2006, 26(2): 191–200 [郝玉江, 王丁, 张先锋. 长江江豚繁殖生物学研究概述. 兽类学报, 2006, 26(2): 191–200]
[10] Kimura S, Akamatsu T, Li SH, et al. Seasonal changes in the local distribution of Yangtze finless porpoises related to fish presence. Marine Mammal Science, 2012, 28(2): 308–324
[11] Kasuya T, Kureha K. The population of finless porpoise in Inland sea of Japan. Scientific Reports of the Whales Research Institute Tokyo, 1979, 31: 1–44
[12] Lacy RC, Pollak JP. Vortex: A stochastic simulation of the extinction process. Version 10.2.14. Chicago Zoological Society. Brookfield, Illinois, USA. 2017
[13] Lacy RC, Borbat M, Pollak JP. Vortex: A stochastic simulation of the extinction process. Version 9.95. Chicago Zoological Society. Brookfield, Illinois, USA, 2009
[14] Lacy RC. Vortex: A computer simulation model for population viability analysis. Wildlife Research, 1993, 20: 45–65
[15] Mei ZG, Chen M, Li YT, et al. Habitat preference of the Yangtze finless porpoise in a minimally disturbed environment. Ecological Modelling, 2017, 353, 47–53
[16] Mei ZG, Zhang XQ, Huang SL, et al. The Yangtze finless porpoise: On an accelerating path to extinction? Biological Conservation, 2014, 172: 117–123
[17] Mei ZG, Hao YJ, Zheng JS, et al. Research progress on the decline mechanism of finless porpoises in the Yangtze River . Chinese Life Sciences, 2011, 5: 519–524 [梅志刚, 郝玉江, 郑劲松, 等. 长江江豚种群衰退机理研究进展. 生命科学, 2011, 5: 519–524]
[18] O’Grady JJ, Brook BW, Reed DH, et al. Realistic levels of inbreeding depression strongly affect extinction risk in wild populations. Biological Conservation, 2006, 133: 42–51
[19] Pei PZ Wang L, Shao YP, et al. Re-introduced Przewalski’s horses’ breeding success and population viability analysis in Anxi National Nature Reserve. Acta Theriologica Sinica of China, 2018, 2: 128–138 [裴鹏祖, 王亮, 邵亚平, 等. 安西极旱荒漠国家级自然保护区重引入普氏野马繁殖成效与种群生存力分析. 兽类学报, 2018, 2: 128–138]
[20] Possingham HP, Davies I, Noble IR, et al. A metapopulation simulation model for assessing the likelihood of plant and animal extinctions. Mathematics and Computers in Simulation, 1992, 33(5–6): 367–372
[21] Ralls K, Ballou JD, Templeton A. Estimates of lethal equivalents and the cost of inbreeding in mammals. Conservation Biology, 1988, 2: 185–193
[22] Shirakihara M, Takemura A, Shirakhara K. Age, growth, and reproduction of the finless porpoise. Nephocaena phocaenoides, in the coastal waters of western Kyuhu, Japan. Marine Mammal Science, 1993, 9(4): 392–406
[23] Tian Y, Wu JG, Kou XJ, et al. Methods and applications of population viability analysis (PVA). Chinese Journal of Applied Ecology, 2011, 22(11): 257–267 [田瑜, 邬建国, 寇晓军, 等. 种群生存力分析(PVA)的方法与应用. 应用生态学报, 2011, 22(11): 257–267]
[24] Xian YJ. Ecological scientific investigation of Yangtze finless porpoise in 2017. Man and the Biosphere, 2018(5): 27 [先义杰. 2017年长江江豚生态科学考察. 人与生物圈, 2018(5): 27]
[25] Yan YB, Chen XJ, Wang JT, et al. Prediction of abundance index of the Humboldt squid (Dosidicus gigas) in the Southeast Pacific Ocean based on a grey system-based model. Progress in Fishery Sciences [闫永斌, 陈新军, 汪金涛, 等. 东南太平洋茎柔鱼资源丰度灰色预测研究. 渔业科学进展. https://doi.org/10.19663/j.issn2095-9869.20190525001]
[26] Yuan Y, Zhang PJ, Wang K, et al. Genome sequence of the freshwater Yangtze finless porpoise. Genes, 2018, 9: 213
[27] Yu DP, Wang J, Yang G, et al. Primary analysis on habitat selection of Yangtze finless porpoise in spring in the section between Hukou and Digang. Acta Theriologica Sinica of China, 2005, 25(3): 302–306 [于道平, 王江, 杨光, 等. 长江湖口至荻港段江豚春季对生境选择的初步分析. 兽类学报, 2005, 25(3): 302–306]
[28] Yang J, Xiao W, Kuang XA, et al. Studies on the distribution, population size and the active regularity of Lipotes vexillifer and Neophcaena phocaenoides. Resources and Environment in the Yangtze Basin, 2000, 9(4): 444–450 [杨健, 肖文, 匡兴安, 等. 洞庭湖、鄱阳湖白鱀豚和长江江豚的生态学研究. 长江流域资源与环境, 2000, 9(4): 444–450]
[29] Yang G, Zhou KY, Gao AL, et al. Study on life table and population dynamics of finless porpoises. Acta zoologica sinica, 1998, 18(1): 1–7 [杨光, 周开亚, 高安利, 等. 江豚生命表和种群动态的研究. 兽类学报, 1998, 18(1): 1–7]
[30] Zhang Y, Li J, Xue YD, et al. Population viability analysis of Sichuan golden snub-nosed monkey in context of climate change in Hubei Province, China. Chinese Journal of Ecology, 2018, 11: 3333–3341 [张宇, 李佳, 薛亚东, 等. 气候变化背景下湖北川金丝猴种群生存力分析. 生态学杂志, 2018, 11: 3333–3341]
[31] Zhang XQ. Population ecology of Yangtze finless porpoise in Dongting Lake and adjacent waters. Wuhan: Institute of Hydrobiology, the Chinese Academy of Sciences, 2011 [张新桥. 洞庭湖及邻近水域长江江豚种群生态学研究. 中科院水生生物研究所, 2011]
[32] Zhang XF, Wang KX. Viability analysis of finless porpoise populations in the Yangtze River. Chinese Acta Ecologica Sinica, 1999, 19(4): 529–533 [张先锋, 王克雄. 长江江豚种群生存力分析. 生态学报, 1999, 19(4): 529–533]
[33] Zhang XF, Wang D, Wang KX. Vortex model and its application on the management of Chinese river dolphin (Lipotes vexilifer) population. Chinese Biodiversity, 1994.2(3): 133–139 [张先锋, 王丁, 王克雄. 漩涡模型及其在白暨豚种群管理中的应用. 生物多样性, 1994, 2(3): 133–139]
[34] Zhang XF. Study on age identification, growth and reproduction of finless porpoises. Chinese Acta Hydrobiologica Sinica, 1992, 16(4): 289–297 [张先锋. 江豚的年龄鉴定、生长和繁殖的研究. 水生生物学报, 1992, 16(4): 289–297]
[35] Zhou, XM, Guang, XM, Sun, D, et al. Population genomics of finless porpoises reveal an incipient cetacean species adapted to freshwater. Nature Communications, 2018, 9: 1276





