
5-HT 是一种广泛存在于脑和消化道的重要的神经递质和胞内信使,在调控机体情绪,体温,睡眠,食欲和代谢活动过程中发挥重要作用。机体90%的5-HT 是由肠道中的肠嗜铬细胞合成、分泌并发挥作用的。近年来研究发现 5-HT 参与调节肠道运动和多种内分泌活动,其含量水平异常可能导致胃肠道内分泌功能紊乱进而导致多种胃肠道疾病的发生,5-HT 受体和转运体表达异常将直接影响肠道 5-HT 含量,间接影响肠道内分泌稳态。本文将详细阐述 5-HT 受体、转运体异常与肠道疾病发生的关系,期望对相关肠道疾病的预防和治疗提供一定的理论依据。
1 5-HT 转运体和受体在肠道分布SERT 是单胺类转运体家族中的一员,是一种负责从突触间隙中重摄取 5-HT 的转运蛋白,其作用的动力来自 Na+/Ka+ATP 酶产生的 Na+的浓度梯度差,是一种高度选择性的Na+和 Cl-依赖性转运体。大鼠、小鼠、牛、猪及人类肠道都有 SERT 存在,SERT 主要表达在肠上皮细胞、产生 5-HT 的神经元及血小板上,其作用可被 5-HT 再摄取抑制剂( SSRIs)所抑制。免疫组化法显示人类、大鼠、豚鼠肠道 SERT主要存在于肠黏膜上皮细胞、肠肌间神经丛,也存在于肠 5-HT 能神经元上。大鼠空肠、回肠、结肠三段肠上皮细胞的顶端和基底细胞膜表面都有 SERT 表达,这与 SERT 在人体小肠分布相一致,但是大部分 EC 不表达 SERT。因此,肠嗜铬细胞( EC) 不能进行 5-HT 灭活。肠道黏膜下层和肠肌间神经丛表达的多巴胺转运体和阳离子转运体也能转运部分 5-HT,但是结合能力和选择性显着低于 SERT。SERT 在肠道内分布广泛,但其活性和表达受多种因素的影响。最近的研究表明促炎因子 IFN-γ 和 TNF-α、腺苷酸、脂多糖均能影响胃肠道 5-HT 能系统,下调肠黏膜上皮细胞 SERT 的表达并损伤 SERT 的活性,导致小肠内细胞外 5-HT 含量升高,引发胃肠道功能紊乱。
5-HT 是唯一通过 13 种 G 蛋白耦联受体和几种配体门控离子通道受体( 5-HT3) 发挥作用的单胺类神经递质。这些受体按结构基础、转导和调控功能不同被分为 7 类,分别是5-HT1( 5-HT1A,5-HT1B,5-HT1D,5-HT1e,5-HT1F) ,5-HT2( 5-HT2A,5-HT2B,5-HT2C) ,5-HT3,5-HT4,5-HT5,5-HT6和 5-HT7,甚至更多的亚型。不同的受体亚型分布部位不同,具有器官分布特性,发挥不同的作用。胃肠道主要靠 5-HT1、5-HT2、5-HT3、5-HT4、5-HT7受体调节胃肠道动力、感觉和分泌功能。
其中,5-HT1A受体分布于中枢及肠神经元上,能够抑制快速兴奋性突触后电位,抑制肠平滑肌动力; 5-HT2A、5-HT2B主要分布于肠肌层,与平滑肌收缩有关,部分 5-HT2A受体分布于黏膜下层和肠肌层胆碱能神经元上,负责调控小肠黏膜上皮的生长; 丁健华等报道5-HT3受体主要分布于肠神经元,目前也被证实存在于肠嗜铬细胞上,能够快速兴奋突触后电位,引起神经递质释放增加,分泌增加; 5-HT4受体主要分布于肠神经元、平滑肌和肠上皮细胞,能使神经递质释放增加,平滑肌收缩,分泌增加; 5-HT7主要分布于平滑肌,肠神经元也有分布,主要功能是松弛平滑肌。
2 5-HT 转运体、受体亚型在肠道中的作用机体几乎 90%的 5-HT 是在胃肠道尤其是在肠 EC 中合成并且分布的,EC 细胞不仅充当黏膜感受传导器而且能够分泌大量 5-HT,对来自管腔的生理和病理性刺激作出应答。5-HT 发挥生理作用后必须灭活,过多的 5-HT 累积导致中毒反应和 5-HT 受体的脱敏。研究表明5-HT 在生理 pH下不能穿过胞膜,必须通过载体转运至胞内灭活,转运过程主要靠肠黏膜上皮细胞膜上的 SERT 来完成,肠道中 5-HT能神经元也可以表达 SERT 将 5-HT 再摄取至细胞内。肠黏膜上皮细胞和 5-HT 能神经元细胞重摄取的 5-HT 在单胺氧化酶的作用下转变成 5-HIAA( 5-羟吲哚乙酸) 而灭活。SERT可以迅速从突触中清除 5-HT,从而减少其在突触间隙中的残留,亦能够有效降低肠道管腔和组织间隙中 5-HT 的浓度。
SERT 毫无疑问是细胞外环境 5-HT 的最有力的调控者,与 5-HT 高亲和力的特性使其发挥严格控制 5-HT 神经递质的强度和持续时间的作用,因此 SERT 的微量变化可引起多种生理功能紊乱。SERT 的重要作用就是控制肠道 5-HT 的含量变化,同时控制 5-HT 受体的脱敏和激活作用。
虽然 5-HT 受体在肠道 5-HT 受体亚型的功能因种群差异和组织器官分布的不同而有所不同,但是目前研究对象主要集中在5-HT1、2、3、4、7几种受体上,尤其以5-HT1A、3、4受体与5-HT 相互作用调节胃肠道运动和分泌活动为主。在中枢神经系统和肠道改变 5-HT 信号通路引起 IBS 病人超敏反应和胃肠道功能损伤,这种自分泌和旁分泌的神经递质及其受体在IBS 症状发生过程中起关键作用,关于 IBS 治疗方案最初的药理学试验主要集中在检测调控肠道功能的受体上,如 5-HT1、5-HT3、5-HT4、5-HT7亚型。研究报道 5-HT1A、5-HT3、5-HT7受体亚型在调节小鼠结肠迁移运动的初始、产生、传播过程中发挥重要作用,下行 5-HT 能神经元的活动对结肠迁移运动是必不可少的。在肠道,5-HT1A受体主要表达在肠神经系统,尤其是在黏膜下层和肠肌间神经丛,负责调控肥大细胞脱颗粒和介质( 包括组胺) 的释放。5-HT3、5-HT4受体拮抗剂能显着抑制胃窦消化间期运动力复合波并显着抑制结肠动力,在结肠近端这种抑制具有浓度依赖性,但是结肠远端抑制作用无浓度依赖性,而 5-HT2B受体拮抗剂对正常的结肠动力无收缩影响。有研究表明 5-HT 通过 5-HT4受体和 cAMP-及 Ca2 +依赖性的信号通路刺激小鼠十二指肠碳酸氢盐的分泌,而 5-HT2受体与 5-HT3受体并未或很少参与小鼠十二指肠碳酸氢盐的分泌调节,5-HTl受体不参与十二指肠碳酸氢盐的分泌调节。对于豚鼠的研究发现,5-HT3受体介导和调控小肠的许多功能如电解液的分泌、肠嗜铬细胞 5-HT 分泌以及小肠平滑肌的收缩和舒张等。Mani-ni等报道一定剂量的 5-HT4受体激动剂维司曲格显着增强小肠和结肠运输,并加速结肠排空。Hoffman等报道 5-HT4受体激动剂维司曲格能够促进结肠推进运动,减弱内脏高敏感性。体外试验证明5-HT7受体在马的肠肌间神经丛上高水平的表达,提示 5-HT7受体可能参与马肠道收缩运动的调节。
3 5-HT 转运体和受体在肠道疾病发生中可能的机制随着 5-HT 信号系统在胃肠道的生理及病理中的作用得到解答,5-HT 受体及其转运体 SERT 在胃肠道的蠕动反射和分泌中的作用也成为胃肠道疾病研究的焦点。SERT 表达异常与精神状态、胃肠道功能状态均紧密相关。IBS 和结肠炎患者黏膜层 SERT 表达均出现下降,提示黏膜炎症可能和SERT 表达减少相关。研究发现SERT 基因敲除的大鼠虽能存活,但是肠道功能异常,大便水分明显增多,少部分大鼠出现便秘及腹泻、便秘交替症状,推测腹泻可能是由于 SERT小鼠 5-HT 的信号通路作用增强,或大量未回收的 5-HT 引起肠蠕动反射和分泌反射增强,便秘可能是大量 5-HT 累积导致 5-HT 受体脱敏引发的短暂性肠运动障碍。肠道 SERT 表达减少,导致 5-HT 累积过度,作用于 5-HT 受体,引起受体的脱敏和表达水平下调,可能是 SERT 参与影响胃肠道功能的机制。免疫印迹显示缺少 5-HT 再吸收转运体的小鼠( SERTKO) 黏膜层的肠壁 caspase-3 活性显着增强,凋亡细胞数量较 WT 小鼠显着增多,但 SERTKO 小鼠是在增加隐窝干细胞增殖的基础上增强绒毛顶端细胞的凋亡保持肠上皮细胞增殖凋亡平衡的。有研究报道SERT 在人类肠上皮样Caco-2 细胞系中的活性和表达受 IL-10 的影响,并且具有浓度依赖性。高浓度时通过 IL-10R 和细胞内 P13K 信号通路诱导细胞膜表面 SERT 活性和表达的升高,IL-10 低浓度时通过独立于 IL-10R 的通路影响转运体的能力而抑制 SERT 的活性。Toll 样受体 3( TLR3) 激活可能通过 p38/MAPK 通路抑制人类肠上皮样 Caco-2 细胞 SERT 的表达和活性,增强细胞外 5-HT 浓度水平导致炎症反应。另外,SERT 基因表达受其上游 5-HTTLPR 区调控,该调控区存在基因多态性,目前 SERT 基因多态性与胃肠道功能性疾病关系的研究主要集中在肠易激综合征上,而且已证实 SERT 基因多态性与IBS 的不同临床症状有关。5-HTTLPR 突变影响 IBS-C( 便秘型肠易激综合征) ,但对 IBS-D( 腹泻型肠易激综合征) 、IBS-M( 混合型肠易激综合征) 无影响,而且这种影响仅存在于东亚人群。
在 5-HT 的 7 个受体亚型中 5-HT2、5-HT3、5-HT4是研究报道最多也是与胃肠道运动及内脏敏感性紧密相关的 4 种受体。关于 5-HT 与受体相互作用的通路主要有神经通路和非神经通路。Day 等报道 5-HT3受体通过非神经元通路介导了鼠结肠末端氯离子的分泌,因 5-HT3受体不仅存在于肠神经元,也存在于嗜铬细胞,用 5-HT3受体拮抗剂进行神经阻抑,同时给予 5-HT3受体激动剂诱导了结肠氯离子的分泌,提示黏膜 5-HT3受体可能通过非神经元通路调节结肠氯离子的分泌。5-HT 还通过 5-HT3受体增加 ICC-肠道起搏器细胞的活性,5-HT3受体通过影响细胞外 Ca2 +的浓度间接影响 ICC 的活性,因此 5-HT3受体可能是肠道功能性运动性失调的研究对象。人类回肠对 5-HT 的分泌反应的非神经性通路主要是由 5-HT4受体介导,而在结肠 5-HT2受体发挥重要作用。因此,5-HT2、5-HT3、5-HT4受体可能通过非神经性通路刺激胃肠蠕动和胃肠道分泌。
5-HT 在肠道发挥作用的神经通路是通过激活神经元上的 5-HT2、5-HT3、5-HT4受体发挥作用的。免疫组化双标和单标发现蟾蜍大肠末端 5-HT 和神经肽( 神经节肽/生长抑素、VIP) 共表达,且共表达的神经元沿 IPANs( 内在的初级传入神经元) 分布。5-HT 通过激活神经元上 5-HT2、5-HT3、5-HT4受体引起神经递质 NO 的释放,进而松弛和收缩豚鼠结肠基部的纵形肌。研究报道 5-HT4受体主要表达在肠神经元上,在肠肌层神经节和黏膜下神经节 5-HT4受体主要表达在 IPANs 上,促分泌神经元几乎不表达,5-HT4 受体激动剂能够作用于 IPANs 和其他肠神经元形成的突触从而增强突触传导的作用。5-HT3受体在内脏疼痛传导通路中起重要作用,自主神经和肠传出神经元均表达 5-HT3受体,刺激这些部位的受体,可导致 Ach 和 SP 释放。因此,阻断 5-HT3受体作用部位可以降低内脏敏感性,并且显着改善 IBS病人的症状。有文献报道,5-HT 在印甲薪导致的小肠溃疡中因受体的不同而扮演双重作用,5-HT3受体通过增加炎症反应使小肠损伤恶化,而 5-HT4受体通过激活乙酰胆碱的释放引发 N 受体效应而发挥抗溃疡效应。然而,新的研究报道神经型 5-HT 可以通过与胆碱能神经元上的 5-HT2A受体结合,激活神经元引起 Ach 的释放至黏膜层,促进黏膜上皮细胞的增殖和转移,进而促进小鼠回肠黏膜上皮的生长,在黏膜下层和肠肌层神经节出现 HuC/D 双标神经元、5-HT2A、ChAT 三重标记活性。几乎所有胆碱能神经元都表达 5-HT2A免疫反应活性,说明神经型 5-HT 通过与 5-HT2A受体结合间接刺激乙酰胆碱神经元释放 Ach,从而促进小鼠肠黏膜上皮细胞的生长。
总之,5-HT 通过结合受体参与调控肠道运动、感觉及分泌活动的机制大概有以下三种: ( 1) 直接作用于黏膜上皮细胞或者平滑肌产生分泌活动,如分布于平滑肌上的 5-HT4、5-HT7受体都是起舒张作用的受体,而 5-HT2A是收缩作用的受体,5-HT 与平滑肌受体作用可直接调控平滑肌的运动。( 2)5-HT 作用于内源性传入神经元( IPANs) 及中间神经元,产生并调节分泌、蠕动反射运动,如肠嗜铬细胞感受肠腔刺激释放 5-HT,通过与 5-HT 受体结合激活黏膜下神经元,黏膜下神经元在释放乙酰胆碱( Ach) 的同时释放降钙素基因相关肽及不同的神经递质如 VIP、NO 等兴奋或抑制胃肠道运动及分泌活动。( 3) 5-HT 还可能作用于外源性传入神经突触后膜上的部分 5-HT 受体,将感觉信号传入中枢神经系统。
4 5-HT 受体在中枢内的分布、表达和功能在中枢神经系统 5-HT 能神经元主要位于中缝核,通过上行和下行通路作用于大脑广泛区域。目前已经确认几种疾病发生时 5-HT 信号系统损伤,包括抑郁、焦虑、精神分裂症、偏头痛和饮食障碍等疾病。在中枢 5-HT 受体被分为 7个亚类 5-HT1A-F,5-HT2A-C,5-HT3,5-HT4,5-HT5,5-HT6,5-HT7。几种受体亚类中除 5-HT3受体是促离子型受体外,其他均是代谢型受体。5-HT1A受体主要分布在 5-HT 能神经元和参与调节情感和认知功能的大脑回路上的非 5-HT 能神经元上。5-HT1B和 5-HT1D受体主要以自受体形式表达在 5-HT能神经元上并且以异受体形式表达在类胆碱能、氨基丁酸能、谷氨酸能神经末梢上,它们消极地耦合到腺苷酸环化酶上并抑制神经递质释放。5-HT2A受体随机地表达在脑部并调控认知过程,也几乎是所有抗精神病药物作用的靶点。在中枢神经系统,5-HT3受体主要表达在部分脑区的神经末梢,突触后神经末梢 5-HT3受体介导快速兴奋性突触传导,而突触前神经末梢 5-HT3受体介导和调控神经递质的释放。5-HT4受体仅表达在突触后细胞上并且刺激腺苷酸环化酶的活性。5-HT7受体主要与小窝蛋白共表达在脂质筏上,5-HT 与 5-HT7受体结合的信号通路受小窝蛋白的调控。
综上所述,机体中 90%5-HT 由肠嗜铬细胞分泌,少量由中枢内 5-HT 能神经元合成。无论在外周还是中枢均由SERT 重摄取、灭活。5-HT 发挥生物学效应主要通过结合细胞或神经元上的 5-HT 受体,包括 GPCR 和 ICCR 两种类型,启动细胞内反应。详细的传导通路汇总如图 1 所示。【1】
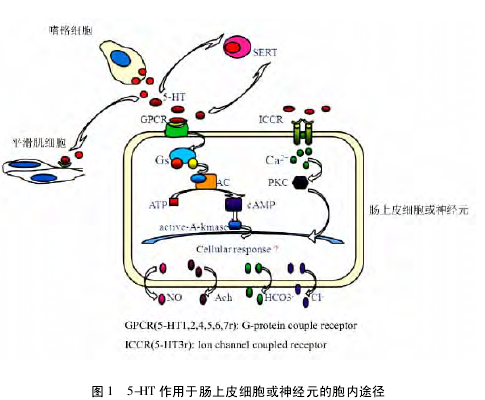
本综述通过详细阐述 5-HT 受体/转运体异常与消化道疾病相互作用关系及可能的机制,期待为 5-HT 相关的肠道相关疾病的预防和治疗提供理论依据。但是,随着 5-HT 亚型的分布和功能研究的深入,肠道发生疾病时具体哪种受体亚型是通过神经元通路还是非神经元通路起主导作用; SERT参与肠道疾病发生具体的发病机制是什么,SERT 基因多态性在与 5-HT 相关的肠道疾病中扮演怎样的角色,都是目前亟待研究的问题。





