����ϸ���쵰�ף����������������ɣ���������������ɵģ�������ģᵰ�ס����������ṹ���о�����鵰�ף��������������ȣ����⣸�������۵��������νṹ���뼡�쵰�ס�Ѫ�쵰�ײ�ͬ��ϸ���쵰����һ������λ���ס������ද���ϸ���쵰�㷺�ֲ��ڸ��࣬���࣬�ԣ��Σ�����Ĥ��ʳ�ܣ����ȸ������١��ڴ������У����ֲ�����άĸϸ���������ϸ����ϸ���ʣ���������Ԫ�У�����ϸ���ʼ�ϸ�����о��зֲ�����ʾ���������о���ij��������������ܡ��ԣ��������귢��ϸ���쵰��������Խ��Խ����о������������ȱ��Ӧ��״̬ʱʱ�����ű�����֯��ϸ������Ҫ��ɫ����ϸ���Ǵ��ԵĻ������ܵ�λ�������Ŵ��Եĸо����˶������䣬˼ά�ȸ����������ܡ��ٴ������ƣȣɣ�ʱ����������ĵ�����Ԫ�Ĵ�������Ͳ��˵�Ԥ���кܴ�Ĺ�����������Ϊֹ������ϸ���쵰��������������ԭ����Ԫȱ��ʱ������о����ޡ���ʵ��ͨ���ң��죭�������УãҼ��ף��������������ϸ���쵰����ԭ����Ԫ��ȱ��ʱ�ı��Ϊ�о�ϸ���쵰����ȱ��Ӧ����ϸ���еĵ���ͨ·�ṩʵ��������Ӷ���һ��Ϊȱ��ȱѪ���Բ��е����Ƶ춨������
���������Ϻͷ���
������������ʵ�鶯���Ҫ�Լ����������У�����ģӣĴ�������ͷ��ѧҽѧԺʵ�鶯�������ṩ���̣��ǣ̣գ��ԣ��ͣɣΣţ��ǣɣ£ãϣ����ģͣţͣ��ƣ������������ǣɣ£ãϣ����£�����Ӫ�����ӣ��ɣΣ֣ɣԣңϣǣţΣ����ԣң٣УӣɣΣ����������� �ţģԣ����ǣɣ£ãϣ����Σţգңϣ£��ӣ��̣��ǣɣ£��ãϣ�����ۣ��ģ������� ���ӣɣǣͣ������ңţ��̣ԣɣͣţ��Уã��Լ��У����������﹤������˾���������������黯�Լ��У��������£���ϩ����øһ�����������£����ÿ���������������£���β���������һ�����人��ʿ�£������쵰��һ�����ӣ��Σԣ��������ö���������ɭ������������������ձ��ϣ��̣٣ͣУգӹ�˾�����ãϣ������䣨���ݾ����豸����ӫ�ⶨ���Уã��ǣ��̣ɣƣţ���ȫ���������̣ţɣã����������ǣ��Ϻ����⣩��ø���ǣ��£����䣩��
����������������
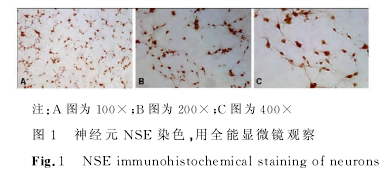
��������������ϸ������������ȡ�У������̥�����Ƥ�㣬�ã����������������������ú�������̥ţѪ��ģģͣţͣ��ƣ����������������Һ����̨����Ⱦɫ����ϸ���������ԣ����������ֲ��Ԥ���ö����������������װ壨�УãҼ�����ӡ�������ã��������װ��ڣ����룸�ã������ڣ����棬�����ãϣ����������н�������������ϸ�����ںΣţգңϣ£��ӣ��� �������£������̣��ǣ̣գԣ��ͣɣΣż���������ÿ�����軻Һ���Σ��������ڣ�������Ԫ������ϩ����ø���Σӣţ�������֯��ѧ������ϸ�����д��ȼ�����
����������������Ԫ���ȼ����������������ã�����ۼ�ȩ�̶����״�˫��ˮ���ݣ���ɽ��Ѫ���գ���һ�����������ã�����棩���ã�°�����ҹ���ڣ���У£ӽ�ϴ�Ӷ�������������������������ø���������У£ӽ�ϴ�μӣģ�����ɫԼ�����������¹۲���ɫ��������У£ӳ�ϴ�����ɣ�������Ƭ�����������ѡȡ��������Ұ������
��������������ϸ��ȱ����ʵ��ǰ�������������ãϣ��ͣ������Σ��������й�ҹ��ʵ��ʱ����ϸ�������Ϊ���飬���У�����Ϊ�����飨���裩�����ࣳ����������ΪԤȱ������������Ȼ����룵���ãϣ��ͣ������Σ����������մ��У��ֱ�ȱ�����裬�����裬�����裬ȱ�����ò����Ǽ���������Ũ�ȣ�����Ũ���ڣ�����������֮��Ľ�����һ��ʵ�顣
��������������ӫ�ⶨ�����Уã��ãԣ�����죨�ԣ����ᣩ���ȷ¡���������������Ҵ��ѽ�ϸ������������ңΣ����ʵ��ģģţУ�ˮ �� ����ңΣ����� �� �� �� �� �� �� �� ���ңΣ�Ũ�Ⱥͣϣ�ֵ������������������������ת¼ø�����£������ңΣ���ת¼�ɣ�ģΣ�������ӫ��Ⱦɫ��������ʵʱ�������ø����Ӧ��������������塡�������壭��������Уãң�����ȡ����걾�����ߣ�����������ã�ֵ����������ϵ ͳ �ģУãҵ� �� Ӧ �� ������ һ �� �Σ����棳���ڶ��Σ����棳�����棳��������ѭ���������Σ����� ���������� ������������ �����Уã�����������¡��ã٣ǣ»�������Ϊ�������ããԣǣǣԣǣ��ǣ��ǣԣԣãԣԣԣǣԣǣ����ã����������Σ����������ã��ǣ����ԣǣ��ã��ããã��ǣ��ǣ��ǣ����ԣã��������� �Σ���β���������� �Σ���������Ϊ���������ãããԣǣ����ǣԣ��ãããã��ԣԣǣ����������Σ����������ԣ��ãǣ��ãã��ǣ��ǣǣã��ԣ��ã��ǣ����������Σ���
������������������ӡ������ӣ���μ�̣���ȥ�ۼ��ѽ�Һ�����Уͣӣƣ������ã��������װ��ڣ����������ñ����ѽ⣳�� ���������������� �������Ĺ��У������£�������������������ģ�����ȡ���岢��װ�ڣ������������Ĺ��У������ڣ�����������б��á��ܵ����ʳ����������ӣģӣ��У��ǣŷ�����ת�����У֣ģ�Ĥ�ϣ��ã�������֬ţ�̷��Һ���·�գ���������˵ģã٣ǣ¶��¡���壨���ã���������β�����������ã����������������������ҹ���ԣ£ӣ�ϴĤ���������×���Σ������������������ø��ǵ��ÿ�����������ã���������������ˮƽҡ���лζ������·������裬�ԣ£ӣ�ϴĤ�������×���Σ������ع⡣�ãѣ�������������ϣ��ͼ�����ϵͳ�Է���ϵͳ�ԣף�����������������з�����
��������������ϸ������ʵ��ÿ��ȱ������������������Ԫ��ȱ��ǰ�������������У£�ϴ����ÿ���ӣããˣ��Լ�������μ�̣������������裬ø���Dz����ϣģ�ֵ��
����ȱ�������������������У£�ϴ����ÿ���ӣããˣ��Լ�������μ�̣������������裬ø���Dz����ϣģ�ֵ������������ԣϣ�ֵ���ϣģ��ϣģ����ϣģ����ֱ��ã��衢�����衢�������룰����Ƚϼ����ȱ�������ϸ���������ʡ�
������������ͳ��ѧ�������������ԣ���±���ʾ������������ϲ��õ����ط�������������Ƚϲ��ạ̃ӣļ��顣�У���������Ϊ������ͳ��ѧ���塣
���������
��������������Ԫ���ȼ�����ϸ��ȱ�����ӣĴ������Ƥ����Ԫ���������� ��������ĺ�����ϸ������״̬���ã����履����������Ρ��Σ������ʶ���Σ������ḻ���˴�����Լ�ɼ���ͻ�������������Ԫ������ϩ����ø���Σӣţ�������֯��ѧ�������ȼ��������ʾ��Ԫ���ȴ��ڣ���������ͼ����ϸ��ȱ�������Ǽ�����Ũ�Ȳ����ڣ������� ����������֮�䡣
����
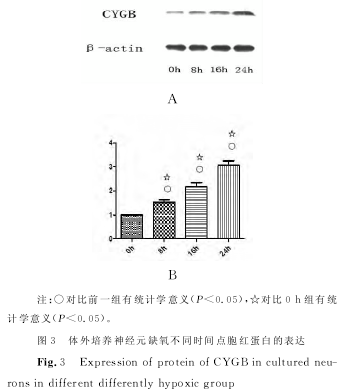
������������ӫ�ⶨ���Уã� ����������飬ȱ����ã٣ǣ��£��ңΣ�����ȱ��ʱ�������ߣ����裬�����裬������ֱ��ǣ�����ģ���������±������������������������±������������������������±����������������ÿ�飨�������֮�������ͳ��ѧ���壨�У���������������ͼ���������ֵ��������
����
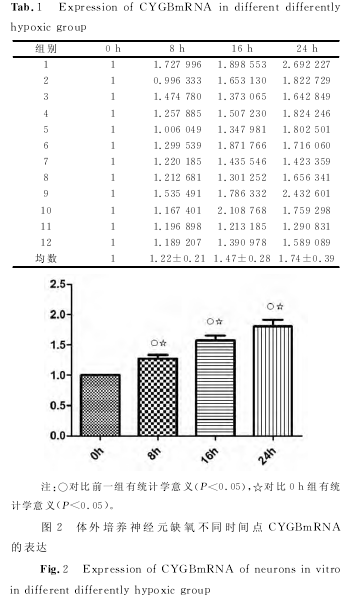
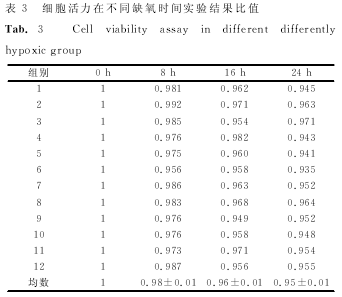
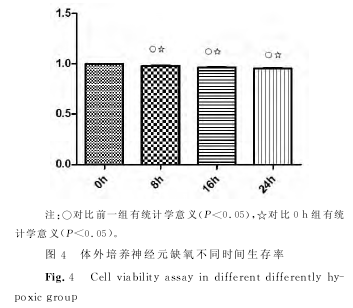
����������������ӡ������ ����������飬ȱ����ã٣ǣ�����ȱ��ʱ�������ߣ����裬�����裬������ֱ��ǣ�����ģ���������±������������������������±������������������������±��������������ÿ�飨�����֮�������ͳ��ѧ���壨�У���������������������ͼ����
����
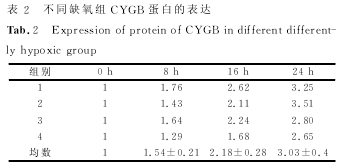
������������ϸ������ʵ���� ����ȱ��ʱ���ӳ��������ʽ��ͣ����裬�����裬������ֱ��ǣ�����ģ���������±������������������������±������������������������±����������������ÿ�飨�������֮�������ͳ��ѧ���壨�У���������������ͼ������������
��������
������ʵ����Ԫ���ȴﵽ���������ϣ���Ũ���ڣ�������������������ȷ���������ȷ�ԡ�ϸ��������������ʾȱ������Ԫ��û�д������������������أ����������У���������ʣ����������������Ԫ���ڴ���һ�ֵ���ѹ�Ļ����йء�������֪����ϸ���Dz�������͵��Ļ��������ȱ������Ԫ�ĸߴ���ʱ�֤�����㹻����ϸ�������ã٣ǣ��£��ңΣ��͵��ף��Ӷ�����ʵ��ӳ�ã٣ǣ�����ϸ��ȱ��ʱ��α����ʵ��ӫ�ⶨ���УãҼ��ף���������������״�֤ʵ�˵���ʹ��ϸ���ں���ˮƽ�͵���ˮƽ��������ã٣ǣ¡�����ǰ����������֯��ϸ��������ʵ����������š��ƣ������֤ʵ����ȱ�����ġ��Ρ����⡢���У��ã٣ǣº������ߡ�������ȱ��ʵ���У����ְ�ϸ������������ϸ����ȣΣ�����֧������Ƥ��ϸ����£ţ��ӣ����⡢������ϸ����ȣ�̣ᡢ���ν���ϸ����ϸ�����еģã٣ǣº��������ߡ����ߵij̶���ȱ����ʱ������س̶ȳ����ȡ���Ȥ���ǣ��ڱ������У�����������飬��������߱����뵰�����߱�������ƽ�У�����ÿ��ʱ�������߱������Ը��ں����飬������뷭����̺ͷ����ļӹ��������йء�ϸ���쵰�ף��ã٣ǣ£����ڸߵȼ������Ϸ��ֵĵ������鵰�ף����㷺�ֲ������������֯���١���ѧ���Ʋ���ͬ�����д������������Ĺ��ܡ�
�����ġ��ȣ���������Ʋ�ȱ�������£ȣ����Σ��ģȵȻ�ԭ�����ʵĴ���ʹ�ã٣ǣ²����乹�������ͷ������Ӷ�ʹ�������Ч����ʹ�������������Ĺ��ܡ����о�֤ʵ�ˣã٣ǣ¾��й�������ø�ͳ��������绯ø�����ã���������������ԭ߲�����л���������� ���ңϣӣ����� Ϊ �� ʹ�ã٣ǣ¹�������������Ӧ���DZ���ϸ����������������ңϣӣ���֬Դ�Ի������ʣ����ͣģΣ������ˣ����ϸ���������ʡ��ó��ã٣ǣ����ʹ�ңϣ�����ٶȼ�����ϸ�����������ߡ�
����Χ����ȱ��ȱѪ���Բ����ȣɣţ�һֱ���������»����²е���Ҫԭ�������������ķ�����ԼΪ��������������������������ķ����ʸ��ߡ��������Dz�����ļ��������Ŵ�������ԣ��������ٻ���ѧϰ�ϰ������ȶ������صĺ���֢��ȱ��ʹ��ϸ�������幦��ʧ����������л�ϰ�����֮�������������ɻ����ңϣӡ�֬�ʹ�������IJ����Ӷ����°��ڵ��ף��ģΣ������ˣ�ϸ���Ļ�������������ˣ���ʱ����������ɻ���֬�ʹ�������ø���Ը��õı�����ϸ������ʮ�����������������˴������о������ǻ�û��һ�ԣȣɣ���֮��Ч�����Ʒ���������һЩ�µ����Ʒ������ڱ��С�
�����ã٣ǣµĹ��ܺ��������ڵĹ㷺�ֲ�ʹ����Ϊ������ȱ��ȱѪ���Բ��ȣɣŵĺ�ѡ��֮һ����ȱ��Ӧ��������£���Ϊ�����ӣã٣ǣ±�����ó��ã٣ǣ·ֱ������ϸ���������ʺͽ���ϸ�����������Լ����ٺ���������֯��������������ǣ�֮ǰ������֮ǰ��ʵ������֤ʵ�ˣã٣ǣ������ȱ��֮��Ĺ�ϵ�����Ǵ�����ϸ������࣬������ֱ��˵����ȱ������£ã٣ǣ�����Ԫ��ֱ�ӹ�ϵ��Ȼ���ٴ������ƣȣɣ�ʱ����������ĵ�������Ԫ�Ĵ�������Ͳ��˵�Ԥ���кܴ�Ĺ����������������Ԫ��ȱ��ʱȷʵ���ϵ��ã٣ǣ£����֤ʵ�ģã٣ǣ���ȱ��ʱ��ϸ���ı������ú�ȱ���������ϸ���������Ʋ�ã٣ǣ���ȱ��Ӧ������ͨ��������ɻ���֬Դ�Ի������ʵ������б�����ϸ������ǿ��Ԫ��ȱ�������������������ϸ��ȱ��ʱ�������ʡ���ˣ��ã٣ǣ»������ܳ�Ϊ���ƣȣɣŵ���;����
����Ȼ�����ã٣ǣ���ȱ��Ӧ���Ĺ�ϵ�����Ĺ����д�����һЩ���顣�ġ��̣�Ƚ���ĸϸ����ϸ����Σ������ڣ������������������ȣ磩�����У���δ �� �֣ã٣ǣ����ߡ��������ʵ����Ƶ���ѹ�����йأ���������²����ද�����Ƥ���ƽ����ѹ�������������������������ȣ磬����ĸϸ��������ϸ����Դ�ġ���ˣ������������ڣΣ���ϸ������ܲ�����“ȱ��”�̼�����ѧ�߽��������ж������ֹ������ģã٣ǣ²�δ���ߡ����������ڼ��ã٣ǣ�ʱ����������ϸ���Ѿ���ȫ���������Ҽ�����ڣ�����֮�������ų���ϸ��δ��ȫ����ʱ�ģ�����֮ �ڣã٣ǣ��Ѿ���ʼ���ߡ�
�����ȣɣ�����ϸ��δ��ȫ��������ˣڣ�������ң����Ľ��۲�����˵���ã٣ǣµı�����ȣɣ��ء�������������ϸ����ȱ��ʱ�����ϵ��ã٣ǣ£����֣ã٣ǣµ��ϵ���������ϸ����һ�����ұ������ƣ���ˣ��ã٣ǣ¿��ܳ�Ϊ���ƣȣɣ�Ԥ���DZ��;��֮һ����Ȼ����кܳ���һ��·Ҫ�ߡ����ȣ���ʵ�鲢δ����ϸ�����ó�������ã٣ǣ����������Ƿ���ľ��б������á���Σ���ʵ�鲢δ̽�����������λ������˽��������û��ơ�������ʵ����Ҫ��һ����֤��Щ���⡣
���������
�����ۣ��ݣˣ�����ᡡ�Σ��ˣ����������ģ£���������ᡡ�ˣ��������죮�ã�������������������桡�ᡡ����������塡����졡������������������������䡡���������ӣԣ��У������������������塡��������������������䡡����������������㡡����������塡������ۣʣݣ��ʣ������졡��桡�£��������졡�ã��������������������������������������������������������������
�����ۣ��ݣӣ����������ȣ��ͣ�����ͣ��ӣ����顡�ȣ��������죮�ӣ������������졡���������������������������������䡡��������ۣʣݣ��ʣ������졡��桡�ͣ����������£������������������������������������������������
�����ۣ��ݣԣ�������ʣԣ��ȣ�������塡�ͣӣ����������������������������������䡡�����������������������塡������������ۣʣݣ��ԣ�塡�ʣ������졡��桡�£���������ã��������������������������������������������������������������
�����ۣ��ݣ£����������ԣ��ţ���£��ף���衡�£��������죮�ã�����������ᡡ������������������塡�����������������������������䡡���������������塡����������ۣʣݣ��ͣ��������£������䡡�ţ������������������������������������������������
�����ۣ��ݣǣ�����ţ��£�����ɣ��ƣ���������ģ��������죮����������������塡���������ۣʣݣ��ԣ�塡�ʣ������졡��桡�£��������졡�ã��������������������������������������������������������������
�����ۣ��ݣȣ������졡�ã����������ǣã��ȣ������졡�ʣ��������죮�������������졡��������������������桡���������������䡡���������������ңΣ������䡡�������������������������塡�������塡������ۣʣݣ��£�����������裬��������������������������������
�����ۣ��ݣͣ�������УУ��ӣ�������ʣͣ��٣塡�ѣ��������죮�ã������������ᡡ�����������������塡�����������������������䡡������塡�����������硡���䡡�ᣭ��������������ۣʣݣ��ԣ�塡�ʣ������졡��桡�ȣ���������������������䡡�ã�������������������ϣ������졡�ʣ������졡��桡����塡�ȣ�������������������ӣ�����������������������������������������������������
�����ۣ��ݣΣ�������顡�ˣ��ϣ��������ᡡ�ȣ��ӣ��������ᡡ�٣��������죮�ã�����������ӣԣ��У������������塡��������������������������㡡������������������������䡡������������������������������ۣʣݣ��̣���������������������������������������������������������������
�����ۣ��ݣӣ����������ͣ��ǣ�����衡�ƣ���������顡�����������죮�ã������������ᡡ�������ᣭ������������������������������塡��������塡���䡡�������������衡����𣭣����������䡡���������������ۣʣݣ��ԣ�塡�ʣ������졡��桡����������졡���������������������������������������������������������
�����ۣ����ݣӣ������������������������顡�٣��ͣ���������ᡡ�ʣ��������죮������������������������桡�����������������������������������������졡����������������������������ۣʣݣ��ȣ�����������������������������������������������
�����ۣ����ݣͣ��ˣΣ��У��������ӣ��ԣ��գ�ˣã��̣���������������䡡���������������������桡�����������������������������������壭����������������������ۣʣݣ��ԣ������������������������������������������������������������
�����ۣ����ݣ̣顡�ңã��̣�塡�ӣˣ��У��������ƣ��������죮�ȣ�������ᡡ������������������������������塡�������������桡���������������䡡��������������������������ۣʣݣ��£�����������裬��������������������������������������
�����ۣ����ݣϣ������㡡�ʣ��ӣ��������顡�ģӣ���塡�̣���������٣��������죮�Σ�������������䡡���������������������������硡������������������졡���������ۣʣݣ��ɣ���������������塡�������������������� ��������졡�������壬������������������������������������������
�����ۣ����ݣƣ����졡�ţ��ǣ�����ţ��ģ������塡�ӣ��������죮�ã������������������������������������䡡�����졡������������������������������������䡡����������������������������������������塡����죭������塡�Уãңۣʣݣ��£�������������䡡�£����������졡�ң������衡�ã�������������������������������������������������������
�ۣ����ݣԣ���ӣƣ��٣��硡�ȣȣ��أ���ģУ��������죮�ͣ����������桡����������������������������������ᣭ���������ᣨ�ȣɣ������������������������𣭣�������������桡�������������ã٣ǣ£����ᡡ���������졡�������������ۣʣݣ��ԣ�塡�ʣ������졡��桡�£��������졡�ã��������������������������������������������������������������
�ۣ����ݣţ���騽���ᡡ�ͣ��ӣ�����ɣ����ԣ�����塡�������������������䡡�����������������������������������ۣʣݣ��ң�����������У����������������������������������������������������
�����ۣ����ݣǣ���أ��У��������ӣ��ԣ��գ�ˣã��ӣ����������桡����塡���������ᣭ��壭������������������������桡��������ã٣ǣ¡������ۣʣݣ��£�������������䡡�£����������졡�ң������衡�ã�������������������������������������������������������
�����ۣ����ݣ£����������ԣ��ǣ�����衡�ƣ��ȣ������ԣ��ң�������������䡡��������������������䡡���������������������������ۣʣݣ���������������������������졡���������塡���䡡��������������������������������������������
�ۣ����ݣȣ������塡�ģ��ˣ���̣��ģ������塡�ӣ��������죮�ԣ�塡���������������塡������塡����졡��������������塡������䡡�������硡��������������桡������������������������䡡�������������ۣʣݣ��ʣ������졡��桡�£��������졡�ã�������������������������������������������������������������
�����ۣ����ݣԣ�������ƣ��ȣ��������ģ����������顡�ƣ��������죮�Σ�������������������������������������졡���������塡��������������ۣʣݣ��ǣ��壬��������������������������������������������
�����ۣ����ݣУ��������ͣǣ��ģ������塡�ӣ��ƣ�������ң���������桡������������������������䡡������������������衡���������塡��������������㡡�����飭�������ۣʣݣ��ʣ������졡��桡���������㡡�������������������������������������������������������������
�����ۣ����ݣȣ����Σʣ��ɣ����������Σ��ģ����ᡡ�ӣ��������죮�ã�������������������������������������塡�ģΣ����������塡����������������������桡����塡������졡����������������������������ۣʣݣ��ͣ������������������������������������������������
�����ۣ����ݣʣ��������֣ͣ��ԣ������ףȣ��ɣ����ᡡ�����������죮�Σ��������������桡���������㣭���������㡡����������������塡�����������硡������ۣʣݣ��У�䣭�������㡡��������裬��������������������������������������
�����ۣ����ݣ���������ǣ��ӣ�����衡�ͣ����ϣ���������衡�ͣţ��������죮�ң��塡��桡���������������������������������䡡������������塡���������䡡����������������������塡������塡��������������桡�������������������ۣʣݣ��Σ��������������������������裬������������������������������������
�����ۣ����ݣ̣顡�ģ��ã���أѣ��̣顡�ףʣ��������죮�ã�������������𣭣����������䡡�������������������������塡������ᡡ�������������塡����塡��������������塡��������ۣʣݣ��Σ�������������졡�ң������裬����������������������������������������.