
随着我国的草坪业蓬勃发展,全国各地进行城市绿化,防治水土流失,建植运动和休息娱乐场地等,对草坪的需求日益增大[1].但是在许多逆境条件下,草坪建植出现困难,草坪草的生长受到抑制,草坪质量和功能受到影响,限制了草坪草的进一步推广应用.盐碱化土壤便是绿化过程中常遇到的自然逆境之一[2].盐碱地在世界范围分布较广,全世界约有9.6×10^5hm²盐碱地,我国有2.7×10^4hm²[3],并随着生态环境的恶化和不合理地开发利用,仍在进一步扩大[4].盐碱地上盐度是影响植物生存、生长和繁殖的重要环境因子[5],在草坪上的具体表现就是造成泛斑、春季发芽晚、秋季早衰、生长不良等,影响草坪的美观,降低草坪的价值[6].因此草坪草耐、抗盐机理的研究是一个重要的课题[1].
植物耐盐性的研究包括盐离子的毒害作用(盐的原初作用)、渗透胁迫和盐离子导致的养分亏缺(盐的次生作用)[7,8].盐分中Na^+和Cl^-对植物的危害较重,极易造成特殊离子效应的单盐毒害,同时对其他离子(如K^+、Ca^2+)的吸收产生拮抗作用,使植株发生营养亏缺,并破坏渗透调节.盐胁迫进一步使得细胞叶绿体和线粒体电子传递泄漏的电子增加,活性氧(ROS)大量积累,造成膜系统氧化损伤[9],从而引起光合电子传递系统失活和光抑制[10],体内激素平衡被破坏和干物质积累下降[11],蛋白质和核酸变性,甚至导致细胞死亡[12].植物可通过限制盐分的过量吸收和调节抗氧化系统清除ROS积累等方式减轻或抵御盐胁迫对细胞的伤害[13].
高羊茅(Festuca arundinacea)作为优良的冷季型草坪草,被广泛应用于各种草坪的建植,是目前我国应用最广泛的冷季型草坪草之一.在高羊茅盐胁迫方面开展了较为广泛的研究,包括重盐碱土上草坪地被植物引种[14]、草坪草耐盐性评价[15-17]、盐胁迫下高羊茅的生理效应[17,18]、高羊茅耐盐突变体筛选[19]、盐胁迫对高羊茅种子发芽的影响[20]、盐胁迫对高羊茅养分吸收与运输的影响[21]以及栽培措施(土壤拌沙、多效唑处理等)对盐胁迫下高羊茅耐盐性的影响[14,22]等.但对于高羊茅遭受盐胁迫后的氧化胁迫反应及其对盐胁迫的耐性缺乏系统的研究.因此,本试验通过用不同浓度NaCl溶液对高羊茅进行处理,研究盐胁迫下高羊茅生长和抗氧化系统的变化特征,揭示高羊茅对盐胁迫响应的生理机制,为冷季型草坪草的逆境生理和抗逆育种提供理论依据,对盐碱地高羊茅草坪管理有较好的指导作用.
1材料与方法
1.1试验材料
供试材料品种名为高羊茅远征.
1.2试验设计
试验在南京农业大学进行,2009年10月在室外采用沙培,将高羊茅种子播种于直径8cm的有孔塑料杯中,Hoagland营养液浇灌,高度保持6cm.
2010年4月,用Hoagland营养液加不同浓度的NaCl处理,使其最终盐处理浓度分别为0,50,100,200,300mmol/L,为了避免盐激作用对幼苗的影响,采用每天递增50mmol/L,直至达到最终盐浓度,以后每天用相应浓度的NaCl溶液浇灌,浇灌量以塑料盆下面的小孔流出为止,每处理3个重复,盐处理期间,进行正常的管理,但不进行修剪,在盐分处理前取样1次,然后分别在达到预定盐分浓度后的第7,14,21天取样.
1.3测定内容与试验方法
生物量:每组中取大小一致的植株,去除枯叶,洗净沙土,吸干植株表面水分,计数茎蘖数,分地上部与地下部称鲜重,然后置于干燥箱中105℃杀青,再于80℃烘干至恒重后分别称干重,生物量以单蘖重(植株总干重/总茎蘖数)表示.
SOD(superoxide dismutase,超氧化物歧化酶)活性测定采用NBT(nitrotetrazolium blue chloride,氯化硝基四氮唑蓝法)法[23],POD(peroxidase,过氧化物酶)活性测定采用愈创木酚法[24],CAT(catalase,过氧化氢酶)活性测定采用Chance法[25],类胡萝卜素含量测定参照张宪政[24]的方法,抗坏血酸(AsA,ascorbic acid)含量测定参照Feibo方法[26].谷胱甘肽(GSH,glutathione)含量测定参照Guri方法,以DTNB[5,5′-dithio bis-(2-nitro-benzoic acid)]显色,在412nm波长下检测[27].MDA含量测定采用赵世杰等[28]改进方法.
1.4数据统计
数据均采用SPSS统计软件分析,并用Excel作图.
2结果与分析
2.1盐胁迫对高羊茅生物量的影响
不同浓度NaCl处理的高羊茅根重呈先上升后下降的趋势(表1).盐处理7d时,在较低的NaCl处理浓度下(≤100mmol/L),根重与同期对照相比略有增加,之后随着胁迫时间的延长呈下降趋势,盐胁迫21d时,各处理均低于同期对照,且随着盐浓度的增加根重下降明显,浓度为300mmol/L的处理生长量最低,为对照的54.93%,其他处理分别降低为同期对照的73.76%,66.23%和63.45%.
茎叶重量短期内逐渐上升,盐处理14d后,50和100mmol/L处理生长趋缓,21d时茎叶干重较对照略低,但差异不显着.200和300mmol/L处理生长量小于低盐处理,14d后呈下降趋势,至21d时与同期对照差异显着,分别降至同期对照的76.66%和68.78%.
2.2盐胁迫对高羊茅抗氧化酶活性的影响
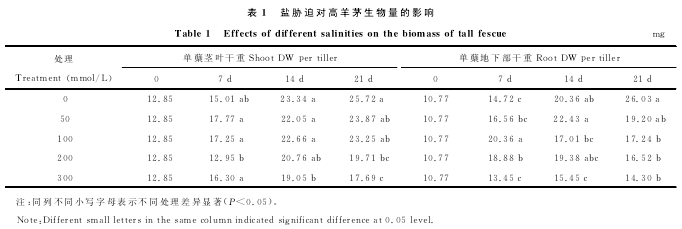
在盐胁迫过程中,SOD活性在前7d逐渐升高直到达到峰值,此期50mmol/L处理的植株酶活性最高,为同期对照的149.35%.之后开始下降,到第21天时,200,300mmol/L处理的植株其活性明显低于同期对照,分别降至同期对照的63.90%和55.76%.50和100mmol/L处理生长在21d时SOD活性较对照略低,但差异不显着(图1).
盐胁迫下CAT活性先上升,第7天时达到峰值,显着高于对照,但盐处理间差异不显着.浓度为100mmol/L的盐处理酶活性最高,高出对照35.31%.之后CAT活性持续下降,第21天时,均低于同期对照,酶活性分别为对照的90.01%,86.97%,74.87%和55.11%.
POD活性在第7天时与对照相比增加幅度不大,300mmol/L的盐处理酶活性低于同期对照,随着盐浓度的增加对POD活性影响越大.第21天时,200和300mmol/L处理的植株酶活性下降幅度较大,分别为同期对照的64.83%和59.14%,50和100mmol/L处理分别为93.47%和85.50%.
2.3盐胁迫对高羊茅非酶抗氧化系统的影响
AsA含量随胁迫时间的延长呈逐渐降低的趋势,21d后,随着盐浓度的增大而明显降低,盐浓度为200和300mmol/L处理的植株AsA含量下降幅度较大,分别比对照降低了9.09%和10.91%(图2).
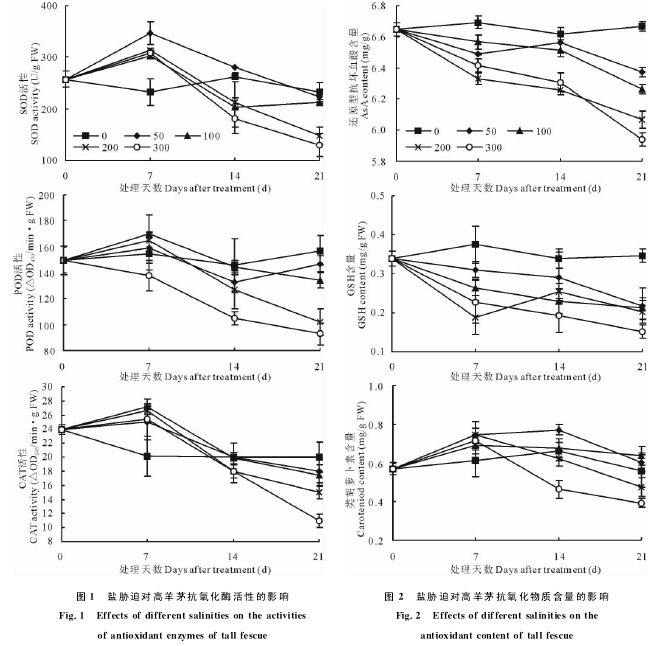
GSH含量随着胁迫时间的延长呈下降趋势,处理浓度越高,下降趋势越明显,第21天时,300mmol/L处理组下降幅度最大,为对照的43.55%,其余盐处理之间差异不明显,为对照的60%左右.类胡萝卜素与抗坏血酸协调作用有利于减少活性氧积累,维持膜的稳定性,各处理的类胡萝卜素含量在前7d均有不同程度的上升,之后逐渐下降,并且盐浓度越高类胡萝卜素含量越低,第21天时,50和100mmol/L处理组与对照差异不显着,200和300mmol/L处理组下降幅度较大,分别为同期对照的85.40%和70.71%.
2.4盐胁迫对高羊茅丙二醛含量的影响
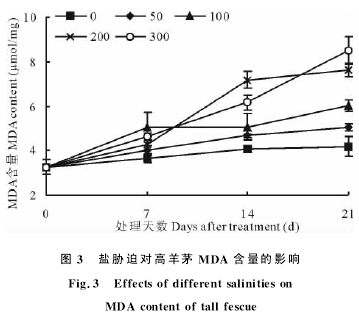
图3盐胁迫对高羊茅MDA含量的影响Fig.3 Effects of different salinities onMDA content of tall fescue丙二醛是膜质过氧化的主要产物,其含量的变化在一定程度上反映了逆境胁迫对植物的伤害,其含量的多少代表了细胞膜脂过氧化水平,反应了植物受胁迫程度的大小.盐胁迫7d时,各处理MDA含量均比对照高,但差异不显着.随着胁迫时间的延长,其含量逐渐升高,第21天时,高浓度(200,300mmol/L)盐胁迫下,MDA含量显着升高,分别比同期对照增加了81.92%和103.43%;中、低浓度(50,100mmol/L)盐胁迫下,分别比对照增加了20.52%和43.19%(图3).
3讨论与结论
生物量是植物对盐胁迫反应的综合体现,即对盐胁迫的综合适应,也是植物耐盐性的直接指标[29,30].试验结果显示短时间内盐胁迫促进了高羊茅根的生长,但随着盐浓度的升高(≥200mmol/L),这种促进作用有下降的趋势.这可能是低NaCl浓度下生长环境能满足幼苗对水分的需求,不会造成盐害,而盐中一定量的无机离子作为渗透调节物质促进了根系生长[31],陈静波等[32]认为盐胁迫下草坪草对光合产物的分配与正常条件下有异,分配到根系的光合产物相对增多,促进了根系的生长.而促进根部的生长,保持良好的根系,有利于植物吸收水分和营养,是植物适应逆境的重要方式.但是随着胁迫时间的延长,植株生物量呈下降的趋势,而且盐胁迫对植株生长的抑制作用随着胁迫浓度的增大而增强,反映了盐胁迫对高羊茅的伤害作用.而且,盐胁迫对地上部与地下部的作用不同,对地下部的影响更大,因此促进根系生长可能是提高高羊茅耐盐性的有效途径.
盐分胁迫对植物的伤害途径之一是氧化胁迫[33],在胁迫初期,植物体内的活性氧清除系统被激活,表现为盐胁迫初期SOD、POD、CAT三种抗氧化酶活性,在不同浓度盐胁迫下均升高,这一结果与张永峰和殷波[34]用不同浓度混合盐碱对苗期紫花苜蓿(Medicago sativa)进行胁迫处理的结果一致,抗氧化酶活性的暂时升高维持了活性氧产生与清除的动态平衡[35].非酶抗氧化物质类胡萝卜素含量有所上升,AsA和GSH含量由于参与自由基的猝灭而呈下降趋势,从而缓解活性氧对植株的伤害作用.但是随着高浓度盐胁迫时间的延长,保护酶系统逐渐被抑制,3种酶活性降低;非酶抗氧化物质含量进一步下降,导致植物总抗氧化能力下降,ROS大量积累,膜脂过氧化作用增强,MDA含量明显上升.细胞内电解质(包含大量植物所需的营养离子)大量外渗,造成植物营养障碍,细胞电化学平衡破坏,代谢紊乱[36],从而影响植物的生长发育,使生物量(尤其是地上部)明显下降.
AsA和GSH是AsA-GSH循环中2种重要的抗氧化剂.通常认为AsA和GSH含量的变化是植物响应盐胁迫最普遍的特征[30],通过试验发现,随着盐浓度的增加和胁迫时间的延长,AsA和GSH含量均呈下降趋势,而且GSH含量的下降幅度比AsA更大.这可能是因为盐胁迫抑制了AsA和GSH的合成或者是盐胁迫下活性氧的大量产生,使AsA和GSH参与了自由基的猝灭所致[37].同时也印证了GSH通过AsA-GSH循环参与AsA的再生,当GSH含量降低时,导致AsA含量降低[38].由此可通过增施外源AsA来保持植株体内AsA浓度,减少GSH的消耗,从而保证植株体内抗氧化剂的数量,以达到提高抗氧化胁迫的能力.其可行性还有待进一步研究.
综上所述,在较低浓度(≤100mmol/L)盐胁迫下,植株通过提高体内的抗氧化保护系统,在一定程度上缓解了盐胁迫的伤害,但是随着盐胁迫时间的延长,超出抗氧化系统的保护范围之后,盐胁迫对植株造成的伤害也随之加重,生长会受到较大影响.
参考文献:
[1]徐胜,李建龙,赵德华.高羊茅的生理生态及其生化特性研究进展[J].草业学报,2004,13(1):58-64.
[2]陈静波,阎君,张婷婷,等.四种暖季型草坪草对长期盐胁迫的生长反应[J].草业学报,2008,17(5):30-36.
[3]李得禄,王继和,李爱德,等.3种驼绒藜属植物种子萌发期耐盐性试验研究[J].中国沙漠,2006,26(6):16-22.
[4]李源,刘贵波,高洪文,等.紫花苜蓿种质耐盐性综合评价及盐胁迫下的生理反应[J].草业学报,2010,19(4):79-86.
[5]肖强,郑海雷,陈瑶.盐度对互花米草生长及脯氨酸、可溶性糖和蛋白质含量的影响[J].生态学杂志,2005,24(4):373-376.
[6]梁慧敏,夏阳,杜峰,等.盐胁迫对两种草坪草抗性生理生化指标影响的研究[J].中国草地,2001,25(3):27-30.
[7]Zheng Q S,Liu L,Liu Y L,et al.Effects of salt and water stresses on osmotic adjustment and osmotica accumulation in a lo-evera seedlings[J].Journal of Plant Physiology and Molecular Biology,2003,29(6):585-588.
[8]Parida A K,Das A B.Salt tolerance and salinity effects on plants:a review[J].Ecotoxicology and Environmental Safety,2005,60:324-349.
[9]Jiang M,Zhang J.Effect of abscisic acid on active oxygen species,antioxidative defense system and oxidative damage in leavesof maize seedlings[J].Plant Cell Physiology,2001,42:1265-1273.
[10]张军,王建波,陈刚,等.Na2CO3胁迫下星星草幼苗叶片电解质外渗率与PSII光能耗散的关系[J].草业学报,2009,18(3):200-206.
[11]刘延吉,张蕾,田晓艳,等.盐胁迫对碱茅幼苗叶片内源激素、NAD激酶及Ca2+-ATPase的效应[J].草业科学,2008,25(4):51-54.
[12]Bethke P C,Jones R L.Cell death of barley aleurone protoplasts is mediated by reactive oxygen species[J].Plant Journal,2001,25:19-29.





