
0 引言
大气电离层相对地面有 360kV 的正电位,导致地球表面存在一个强度约为 130V/m 的自然电场环境.地球上生物的生命活动都受到该电场环境的影响,并在长期的进化过程中逐渐适应了自然电场环境,演化出与之对应的生理特征.当电场环境发生改变,如人为外加高压静电场,必然对其中生物的生命活动产生影响.这种影响被称为高压静电场的生物效应,该效应引起了科学界的极大关注,并由此发展的高压静电技术已经广泛应用到农业、医学等各个领域.如高压静电场可明显延长蔬果的储藏时间[1],提前或迟滞植物开花时间[2];促进植物愈伤组织的生长与分化[3];提高种子发芽率等[4].然而,关于高压静电场对植物叶绿素荧光参数的影响的研究较少.研究高压静电场处理对植物叶绿素荧光参数的影响,不仅对高压静电植物学效应机理研究具有重大意义,而且对于生产实践具有非常重要的指导意义.
叶绿素荧光技术在测定叶片光合作用过程中叶片对光能的吸收、传递、分配、耗散等方面具有独特作用.相对传统气体交换指标,叶绿素荧光参数可以更好反应植物叶片的内在特征.因此,叶绿素荧光技术被视为研究植物光合作用与环境关系的内在探针[5 - 8].本文以"津优 1 号"黄瓜为研究对象,研究了高压静电场处理对黄瓜幼苗叶绿素荧光参数的影响,旨在进一步探索高压静电促进植物生长机理,并为其在种植业中的实际应用提供理论依据.
1 材料与方法
1. 1 供试材料
供试黄瓜(Cucumis sativus L. ) 种子选用津优一号.选取饱满整齐一致的种子,用 5% 福尔马林浸泡种子 0. 5h,洗净后置于蒸馏水中浸泡 24h(置于 28°C恒温光照培养箱内)催芽;将发芽的种子播于装有珍珠岩的穴盘内,温室昼温(28 ±2)℃,夜温(16 ±2)℃,自然光照并浇灌 Hoagland 营养液(pH 6. 5);待培养至第 30 天待用.
1. 2 实验设备
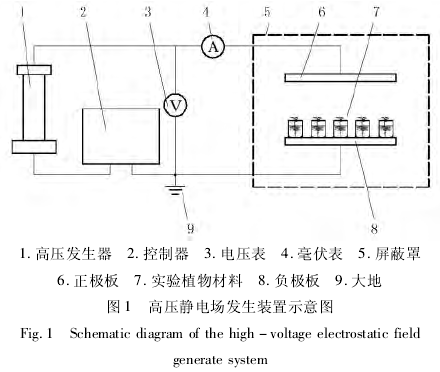
高压静电发生装置如图 1 所示.装置主要由电极板、屏蔽罩与一个直流高压发生器构成(BBG 脉宽调制型直流高压发生器,北京机电研究院高电压技术公司).
1. 3 试验处理
从黄瓜幼苗中挑选大小相近、长势相似的植株,每 10 株为一组均匀放置于不同强度高压静电场下处理.处理组电场强度分别为 0. 5、1. 0、1. 5、2. 0、2. 5、3. 0kV / cm,处理时间 120min.另设一组作为对照(ck),除不加电场外,其余条件相同.整个实验在人工气候室进行,温度(25 ± 1) °C,相对湿度 60% ~70%,光照强度 200μmol/(m²·s).
1. 4 测定方法方法
处理后,使用 IMAGE - PAM 叶绿素荧光成像系统(德国 WALZ 公司生产)测定各株幼苗从植株顶部向下第一完全展开功能叶的叶绿素荧光参数.将待测材料充分暗适应 30min 以上,在室温(25 ± 1 )°C 下测定幼苗的叶片固定荧光 ( Fo)、光化学淬灭系数(qP)、PSⅡ最大光化学效率(Fv/Fm)、表观光电子传递速率(ETR)、非光化学萃灭系数 NPQ 与 qN.
2 结果与分析
固定荧光又称初始荧光,指 PSII 反应中心处于完全开放时的荧光产量[9 - 10].如图 2 所示,当外加高压静电场强度较低时 Fo 基本与对照组维持同一水平.
场强超过 1. 0kV/cm 时 Fo 值开始随外加电场增强而增加,并在 2. 0kV/cm 时达到最大,此后保持在高水平.
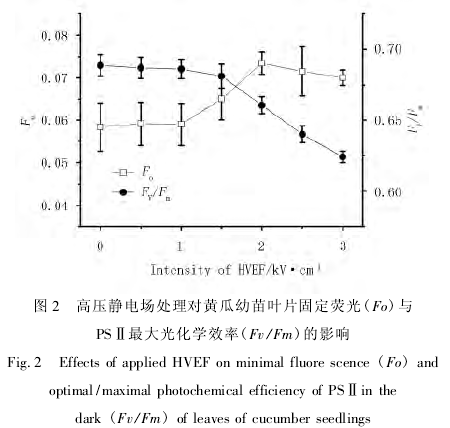
PSⅡ最大光化学效率 Fv / Fm,反映 PSⅡ光化学效率的变化,降低说明其发生光抑制的程度越高[11].如图 2 所示,1. 5kV/cm 以内的高压静电场对黄瓜幼苗光化学效率影响不大,2. 0kV/cm 及以上强度的外加电场使 Fv/Fm 显着降低.
ETR 即表观光合电子传递速率,反映光化学反应的启动速率[10 - 12].如图 3 所示,外加电场较弱时 ETR与电场强度呈正相关,超过 1. 5kV/cm 的高压静电场则使 ETR 值降低,且场强为 3. 0kV/cm 时低于对照组.
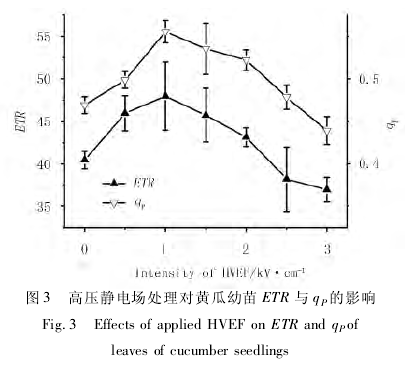
光化学淬灭系数 qP表示光合活性的高低,其随外加电场强度增加的变化规律与 ETR 相似,也呈现"先增后减"的趋势,且于 1. 0kV/cm 场强时取得最大值0. 56,如图 3 所示.
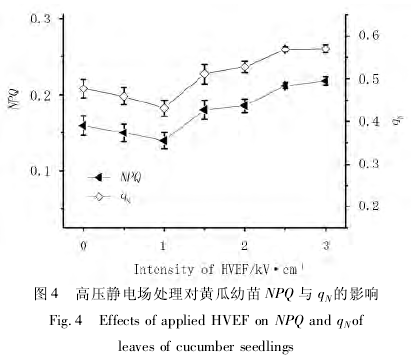
NPQ 与 qN均反映植物细胞的非光化学猝灭状况,即 PSⅡ天线色素吸收的多余光能无法用于光化学电子传递,而以热能的形式耗散掉,对光合机构起自我保护作用[8,13 -15].由图 4 可见,适宜强度的高压静电场处理可降低 NPQ 与 qN值,提高黄瓜幼苗的光能利用率.其中,0 ~ 1. 0kV/cm 以内,NPQ 与 qN随场强增加而降低;反之,电场强度超过 1. 0kV/cm,NPQ 与 qN则与场强呈正相关,耗散的光能增加.
3 讨论
叶绿体间质中悬浮有由膜囊构成的类囊体,类囊体膜上含有光合色素和电子传递链组分,是光能向化学能转化的关键[16 - 19].当类囊体电压处于正常状态下( -70 ~ +165mV)时,荧光产额与类囊体电压呈线性正相关.即,类囊体电压每增加 100mV,荧光产额增加 9% ( ± 2% )[20].当外加电场强度超过临界范围,类囊体电压每增加 100mV,原初电荷分离常数与基本电荷复合速率常数分别下降约 8% 与 50%[21].本研究中,类囊体直径约 0. 5μm,场强增加每1. 0kV / cm ,在类囊体两端产生约为 50mV 的电势差,因而在一定范围内,荧光产额增加.其表现在,随外加电场增强 ETR 即表观光合电子传递速率与光化学淬灭系数 qP升高,电场提高了 PSⅡ的电子传递活性;同时,反映植物细胞的非光化学猝灭状况的参数 NPQ 与 qN下降,以热能的形式耗散掉的光能降低[12,16].因而,黄瓜幼苗对光能的利用率升高,耗散的光能降低,有利于光合作用.高压静电场强度大于为 1. 0 kV/cm时,类囊体电压升至正常状态上限(165 mV),对光能转化的促进作用达到最强.当外加电场继续增强,上述促进随外加电场增强逐渐减弱.超过 2. 0kV/cm 的高压静电场使参数 Fo 升高,表明 PSⅡ反应中心受到破坏或不可逆失活[13,20 -21,24].此外,NPQ 与 qN显着上升,ETR 与 qP下降,说明叶片吸收的光能耗散增多,光能转化效能降低,从侧面反应了光合作用结构遭到破坏[24 - 25].
4 结论
本文对不同强度高压静电场对黄瓜幼苗叶绿素荧光参数的影响进行了实验研究.结果表明,较低强度的外加高压静电场可以促进黄瓜幼苗叶片将光能转化为化学能.其中,在 0. 5 ~1. 5kV/cm 阶段促进作用显着,过高强度的外加电场则不利于光能的转化.该研究为利用高压静电场提高植物对光照利用效能提供了理论依据,并为静电除虫等技术提供了安全电压的数据依据.
参考文献:
[1] Knorr D. Novel approaches in food - processing technology:new technologies for preserving foods and modifying function[J]. Current Opinion in Biotechnology,1999,10(5): 485- 491.
[2] 邓鸿模,虞锦岚,周艾民. 高压静电场促进植物生长技术的研究[J]. 物理,2000,29(9): 550 -552.
[3] 李一. 高压静电场提高顽拗型籼稻花培效率的研究[J].作物研究,1997,11(1): 12 -13.
[4] Moon J D,Chung H S. Acceleration of germination of tomatoseed by applying AC electric and magnetic fields[J]. Journalof electrostatics,2000,48(2): 103 - 114.
[5] Govindjee. Sixty - three years since Kautsky:Chlorophyll afluorescence[J]. Australian Journal of Plant Physiology,1995,22(2): 131 - 160.
[6] Schweiger J,Lang M,Lichtenthaler H K. Differences in flu-orescence excitation spectra of leaves between stressed andnon - stressed plants[J]. Journal of Plant Physiology,1996,148(5): 536 - 547.
[7] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报,1999,16(4): 444 -448.
[8] 徐德聪,吕芳德,刘小阳,等. 叶绿素荧光测定技术的研究[J]. 安徽农业科学,2008,35(35): 11335 -11337.
[9] Kooten O,Snel J F H. The use of chlorophyll fluorescencenomenclature in plant stress physiology[J]. PhotosynthesisResearch,1990,25(3): 147 - 150.
[10] 陈建明,俞晓平,程家安. 叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J]. 浙江农业学报,2006,18(1) : 51 - 55.
[11] 徐晓昀,郁继华,颉建明. 高温胁迫对黄瓜叶片叶绿素荧光猝灭的效应[J]. 甘肃农业大学学报,2007,42(2) : 60 - 62.
[12] 贺立红,贺立静,梁红. 银杏不同品种叶绿素荧光参数的比较[J]. 华南农业大学学报,2006,27(4): 43 -46.
[13] Arnold W,Azzi J. The mechanism of delayed light produc-tion by photosynthetic organism and new effect of electricfields on chloroplasts[J]. Photochemistry and photobiolo-gy,1971,14(3): 233 - 240.
[14] 周秀杰,赵红波,马成仓. 硅对严重干旱胁迫下黄瓜幼苗叶绿素荧光参数的影响[J]. 华北农学报,2008,22(5) : 79 - 81.
[15] Genty B,Briantais J M,Baker N R. The relationship be-tween the quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescence[J]. Biochimicaet Biophysica Acta ( BBA) - General Subjects,1989,990(1): 87 - 92.
[16] 王可玢,许春辉. 水分胁迫对小麦旗叶某些体内叶绿素a 荧光参数的影响[J]. 生物物理学报,1997,13 (2):273 - 278.
[17] 李功藩,蔡琬平,吴亚君,等. 叶绿体结构状态与光化学活性的关系[J]. 植物生理学报,1987,13(3): 295- 301.
[18] 刁丰秋,章文华. 盐胁迫对大麦叶片类囊体膜组成和功能的影响[J]. 植物生理学报 ( ISSN0257 - 4829),1997,23(2): 105 - 110.





