��¡�뿹TYLCV�йصĻ����뷬�ѱ������
ʱ�䣺2014-06-27 ��Դ��δ֪ ���ߣ�ɵɵ���� ����������6353��
�������ѣ�Solanum lycopersicum L.�������з��ѻƻ���Ҷ����Ϊ���������أ�Czosnek & Ghanim��2011�������ѻƻ���Ҷ������Tomato yellow leaf curl virus����� TYLCV���ǵ�����״ DNA ������������ �ȣ�1998������Ҫ�����̷�ʭ���д�����Stansly et al.��2012�����봫ͳ������ȣ������������ڶ̣������ֹ����в��������������ũ����״�Ļ����ڷ��ѿ������������е�Ӧ�������ܵ����ӡ�ͨ����¡�뿹 TYLCV �йصĻ�����ֲ�����ڣ��������ֲ��Ŀ����ԡ�
�����о�������R ����Ը��ֲ�ԭ�������IJ��������п��ԣ�����ֲ��������зֲ��㷺��Flor��1971���������ѿ�¡�� R ����������ĵ����ʱ��ؽṹ������ϸ���е�λ�ã����Խ� R �����ΪNBS-LRR �ࡢLRR-TM �ࡢSTK �ࡢLRR-TM-STK ��� CC �������Hammond-Kosack & Jones��1997����NBS-LRR ���������֪�� R �������������һ�ࣨDangl & Jones��2001�����Ƚϻ������������ֲ���������Ա�����ʮ�������ٸ� NBS-LRR ������ľ�Ϻͻƹϵ� NBS-LRR ������50 ������ˮ������ 653 ����Shang et al.��2009�����ڲ�����Ⱦʱ��NBS-LRR ��������ֲ����ͷ�Ӧ���Թ����Կ��Է�Ӧ�ͼ��˿��Է�Ӧ������ʽ����������Ⱦ��Hull��2002�������ڼ��˿����в�����ֳ�������ڵ�ϸ��ˮƽ��ͨ���ֲ���������ʹ���������ڳ�ʼ��Ⱦλ��������Lukhovitskaya et al.��2005����NBS-LRR �������� NBS-LRR �����к�������λ�㣨NBS�����������ظ�����LRR�������� NBS �������ź�ת���У����и߶ȱ������ϸ�����Ķ�����ܻ���Tan &Wu��2012����LRR Ϊ�߶����Ľṹ������Ϊ������벡ԭ���ʶ����ź�ת�����伤ø�ṹ���ڼ���������ת��Ԫ���з������ã�Jones & Dangl��2006����Ŀǰ��֪ LRR �ṹ��ͨ������ NBS �ṹ��ķ���״̬�����ӿ������ܣ�Takken et al.��2006����
�������ڷ�����Ӧ TYLCV ��Ⱦ��ת¼����������ǰ�ڷ����� 1 �� NBS-LRR ��������ڿ������Ѳ������ϵ������ͨ�� VIGS ���� TYLCV ��Ⱦ�������Ѳ���‘CLN2777A’��û���������ı仯�����û�����һ���Ŀ������ܣ�Chen et al.��2013�����������н�һ����¡�� NBS-LRR ������� ClNLR��ͨ��ӫ�ⶨ�� PCR ���� ClNLR �ڷ���‘CLN2777A’����ҶƬ�и�ˮƽ�����ͨ�������з��Ѳ��Ͻ��� TYLCV �� ClNLR �������ĶԱȺ��ڱ������е�˲ʱ�����һ��֤���˸û����뷬�ѿ�������ء�ͬʱ�ٴ�ͨ�� VIGS ����ԽӲ�ǰ���ѿ�������‘CLN2777A’�ı��ͽ����˹۲죬��ͨ��ӫ�ⶨ�� PCR ��֤��ǰ�ڵ���������
����1�� �����뷽��
����1.1 �������
������ TYLCV ���Ѳ���‘CLN2777A’�� TYLCV ���Ѳ���‘TMXA48-4-0’�ɽ���ʡũҵ��ѧԺ�߲��о����ṩ��Я��TYLCV���̷�ʭΪ���������ڷ������ҷ����ϵ�������Ⱥ��pTRV1��pTRV2�� pTRV2-PDS �������廪��ѧ�����ֽ����������˾� DH5α������ũ�˾� LBA4404��GV3101������ pBI121 ��Ϊ��ʵ���ұ��档������������ CaMV35S �ɱ�ʵ������ pBI121 �Ļ����ϸ�����ɡ�
����TA ��¡���� PGEM-T ���塢����������ø��Xba��BamH��Sac��T4DNA ����ø��DNA �ۺ�ø��DNA �������������� TaKaRa ��˾�������Լ�Ϊ�����������Լ���TIANamp Genomic DNAKit��ѪҺ/ϸ��/��֯������ DNA ��ȡ�Լ��й��� TIANGEN ���﹫˾��������ȡ�Լ��С�PCR ����Լ��С�DNA Ƭ�ο��ٴ���/�����Լ��й��� AXYGEN ��˾������������ Primer 5.0 ������������������ɣ���ί���Ϻ�Ӣ����˾�ϳɡ�
����1.2 ClNLR �Ŀ�¡�����з���
������ǰ�ڵ��о��У���‘CLN2777A’��‘TMXA48-4-0’���Ѳ��Ͻ��� TYLCV ǰ���ת¼�飨NCBIsra ���ݿ��¼�� SRP028618���з��� 1 ����Ӧ TYLCV ��Ⱦ���ϵ����ע��Ϊ���� NBS-LRR �ṹ��Ŀ��Ի���Solyc05g009760.1��ͨ�������Ĺ�����֤�������û�����һ���Ŀ������ܣ�Chen et al.��2013����
�������� Solyc05g009760.1 �������У��� Primer 5.0 ��������������������ȫ������������ P3��5′-TTCACAAGACTTTAGATGTGGCAT-3′�� P4��5′-AGTAGTTCTTGGGGATATGAAGTTG-3′���Կ�������‘CLN2777A’cDNA Ϊģ�壬���� PrimerSTAR �߱���ø��Ӧ��ϵ�����û������С�PCR ��Ӧ����Ϊ��94 ��Ԥ���� 3 min��94 �� 30 s��58 �� 40 s��72 �� 2 min 30 s��35 ��ѭ����72 �� 10 min��
���������յ� PCR ���������� PGEM-T �����ת���˾� DH5α��ת���Ӿ����� PCR ����Ϊ���Կ�¡�������Ϻ�Ӣ����˾��������õĻ�����������Ϊ ClNLR �������������� Conserved Domains�������ؽṹ��
����1.3 ClNLR �� TYLCV �յ��µı������
���������Ѳ���‘CLN2777A’��‘TMXA48-4-0’��Я�� TYLCV ���̷�ʭ���� TYLCV ���� 5 d����ȡҶƬ RNA��M-MLV Reverse Transcriptas��eMBI ��˾����ת¼�ϳ� cDNA���� Actin��AB199316��Ϊ�ڲΣ�ӫ�ⶨ�� PCR ��� ClNLR ����ı��������ClNLR �� Actin ����ֱ�Ϊ ClNLR-forward��5′-CTTTGCGGGTTCGTTCATCTTAT-3′��ClNLR-reward��5′-CGTTTATGTCCACATGCCTCAAC-3′��Actin-forward��5′-TGGTCGGAATGGGACAGAAG-3′��Actin-reward��5′-CTCAGTCAGGAGAACAGGGT-3′��ӫ�ⶨ�� PCR ��Ӧ��ϵΪ 20 μL��2× SYBR Premix Ex Taq?��10 μL������� 1 μL��cDNAģ�� 1 μL������ˮ 7 μL����ϼ�����PCR ��Ӧ����Ϊ 96 ��Ԥ���� 1 min��95 ����� 15 s��60 ���˻� 15 s��72 ������ 45 s��40 ��ѭ������������ 3 �������ظ���
����1.4 ClNLR ����֯�������
������ȡ���ѿ�������‘CLN2777A’��������ҶƬ�����ٺ�ʵ RNA��ÿ����Ʒȡ 500 ng RNA����ת¼�� cDNA ��ϡ�� 1 000 ����ȡ 2 μL ����ӫ�ⶨ�� PCR ��Ӧ��������������ڲ����ӫ�ⶨ�� PCR �ķ�Ӧ��ϵ�ͷ�Ӧ������� 1.3 �ķ������������ 3 ������ѧ�ظ���
����1.5 ClNLR �ڱ����̵�˲ʱ����
�����Ի�õ�ClNLR ����Ϊģ�壬ͨ�� PCR ������ClNLR������������BamH���Sac��ø��λ�㡣PCR ����Ϊ P5��5′-CATGGATCCGGCATTGTTAATCATTAGAGTATG-3′������ BamH��ø��λ�㣩�� P6��5′-ACTGAGCTCTTAGTAGTTCTTGGGGATATGAAG-3′������ Sac��ø��λ�㣩��ֲ��������� pBI121 �� PCR ����ֱ��� BamH��� Sac��˫ø�У��� 16 �����ӹ�ҹ��ȡ 5 μL ���Ӳ���ת���˾� DH5α���ڹ��� LB������ 50 mg · L-1����ù�أ��������� 37 ��������ҹ������¡��ȡ���� DNA ����ø����֤������Ϊ pBI121-ClNLR��pBI121-ClNLR ����ͨ�����ڷ�ת��ũ�˾�LBA4404��
����������pBI121-ClNLR��ũ�˾�������Һ��LB������50 mg · L-1����ù�غ�25 mg · L-1����ƽ����28 ��������ҹ��ȡ����Һ���������壬���������ڻ���Һ��10 mmol · L-1MES��10 mmol · L-1MgCl2��200 μmol · L-1��������ͪ���ڣ����� OD600ֵ�� 0.5��������ͷ 1 mL ��ע������ũ�˾�����Һѹ��5 ~ 6Ƭ��Ҷ�ڵı����̵İ��ҶƬ����һ���ҶƬע����ͬ��������õ�pBI121-GFP��Ϊ���ա�
����ũ�˾�ע�����̲����������У�22 �棬75%����ʪ�ȣ��ڰ������� 2 d��ת���˹�������������28 �棬16 h/8 h �������ڣ����۲���ͱ仯��
����1.6 VIGS ��Ĭ ClNLR ����
������pBI121-ClNLRΪģ�壬PCR��¡��283 bp��ClNLR����Ƭ�Σ�P7��5′-GTATCTAGAGTGTTGTATGTCCGTGGCTGAC-3′�� P8��5′-CATGGATCCGAGAACCCAATGAC TCCCTGC-3′������ Xba���BamH��ø�� PCR ����Ͳ������� pTRV2��Ȼ�û���Ƭ�ο�¡�� pTRV2 �У����� ClNLR ����ij�Ĭ���� pTRV2-ClNLR��
�������� pTRV1��pTRV2��pTRV2-PDS �� pTRV2-ClNLR ���ö��ڷ�ת��ũ�˾� GV3101����ת���ɹ���ũ�˾� 28 ���ҹ����������������������ڻ���Һ��10 mmol · L-1MES��10 mmol · L-1MgCl2��200 μmol · L-1��������ͪ���У����� OD600ֵ�� 2.0�����·��� 4 ~ 6 h ������ pTRV2-ClNLR��pTRV2-PDS �� pTRV2 �������ũ�˾�����Һ�ֱ��뺬�� pTRV1 ��ũ�˾�����Һ�� 1�U1 �ı�����ϣ�������ͷ 1 mL ��ע������ũ�˾�����Һע�뿹�����Ѳ���‘CLN2777A’��Ҷ���������Ҷ�С����У�ע�� pTRV1 + pTRV2 �������ֲ��Ϊ���Զ��գ�ע�� pTRV1 + pTRV2-PDS ��Ϊ���Զ��գ���������һֱ�� 22 ~ 25 �桢���ʪ�� 50%��16 h/8 h �Ĺ���������������
������ע�� pTRV1 + pTRV-PDS �ķ���ҶƬ�ϳ��ֹ�Ư������ʱ����ȡ pTRV1 + pTRV2-ClNLR �ķ����϶���Ҷ�� RNA������ oligo��dT���ϳɵ�һ�� cDNA��Ȼ��ͨ��ӫ�ⶨ�� PCR ��� ClNLR ����ij�ĬЧ�����������P��Ӧ�������� 1.3��
����1.7 ClNLR �����Ĭ�Է������� TYLCV ������Ӱ��
����ѡȡ ClNLR �����Ĭ�ķ���ֲ���ע�� pTRV1 + pTRV2 ������ķ���ֲ��ͬʱ�����ܱյķ������ң��̷�ʭ���� TYLCV �� 7 d ʱ���� CTAB ����ȡ����ҶƬ�� DNA���� SHIMADZU ��˾��BioSpec-nano/230v �ͺŵ����ֹ��ȼƲⶨ�� DNA Ũ�Ⱥ���ϡ��Ϊ��Ũ�� 300 ng · μL-1���á�ӫ�ⶨ�� PCR ��ⷬ��ֲ�����ڵ� TYLCV ���ۣ��� Actin Ϊ�ڲΡ����м�� TYLCV ��������������Ϊ TY-AV494��5′-GCCYATRTAYAGRAAGCCMAG-3′�� TY-COPR��5′-GANGSATGHGTRCADGCCATATA-3′��Actin ���Pӫ�ⶨ�� PCR �ķ�Ӧ�������� 1.3��
����2�� ��������
����2.1 ClNLR ����Ŀ�¡
��������ǰ��ͨ���ȽϷ��Ѳ���‘CLN2777A’��‘TMXA48-4-0’���� TYLCV ǰ���ת¼�飨NCBIsra ���ݿ��¼�� SRP028618���������ڽ��� TYLCV ��1 �� NBS-LRR ��Ի���Solyc05g009760.1���ڿ�������‘CLN2777A’���ϵ�������ڸв�����‘TMXA48-4-0’���������Ա仯��Chen et al.��2013����
�����������н�һ��ͨ�� PCR ��¡�û����ڸû����ע����Ϊ NBS-LRR ��������֮����Ϊ ClNLR���ṹ�������� ClNLR ������뵰���� N �˵� 43 ~ 323 λ������Ϊ���ؽṹ�� NB-ARC��480 ~721 λ����������Ϊ���͵� LRR �ṹ��ͼ 1����
����
����2.2 ClNLR ����ı������
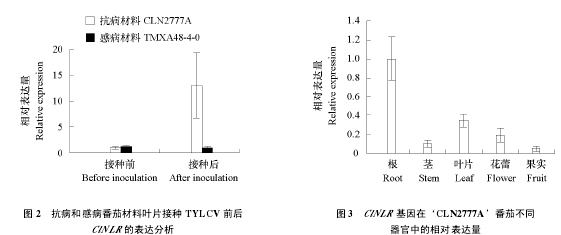
����Ϊ����֤ ClNLR �����ת¼�������������ӫ�ⶨ�� PCR ��� ClNLR ��Ӧ TYLCV ��Ⱦ�ı���������������Ѳ���‘CLN2777A’�в����Ѳ���‘TMXA48-4-0’ͨ���̷�ʭ���� TYLCV��5 d �� ClNLR �ڿ��������б�������Ƚ���ǰ�ϵ���Լ 12 �������ڸв��������������Ա仯��ͼ 2�����������仯������ת¼������Chen et al.��2013������һ�¡�ӫ�ⶨ�� PCR ��һ����� ClNLR �����ڿ�������‘CLN2777A’�е���֯������������� ClNLR�ڸ���������ǿ�����ΪҶƬ���ڹ�ʵ�б���������ͼ 3����
����
����2.3 ClNLR ˲ʱ�����������̹����Է�Ӧ
���������� ClNLR ����ֲ����������ũ�˾�ͨ��ũ�˾�����ע�䱾����ҶƬ 15 d ��ע�� ClNLR �����һ��ҶƬ�ϳ����˻����ߣ���ע�� GFP �������һ��ҶƬû���κα仯��ͼ 4�������� ClNLR �����յ��˱����̹����Է�Ӧ��������ֲ���������Ӧ��ء�

����2.4 ��Ĭ ClNLR ����Ӱ�췬�� TYLCV ����
����Ϊ���о�ClNLR�ڷ����п�TYLCV�����ã���������ͨ��TRV��Ĭ�������ѵ�ClNLR����������ע�� TRV ��Ĭ���� 15 ~ 20 d��ע�� pTRV1 + pTRV2-PDS �����Զ���ֲ����Ҷ���ֹ�Ư����������ʱ�����ƣ�ҶƬ���ֹ�Ư�����������ԣ�ͼ 5�������� VIGS ��Ĭ�ɹ���

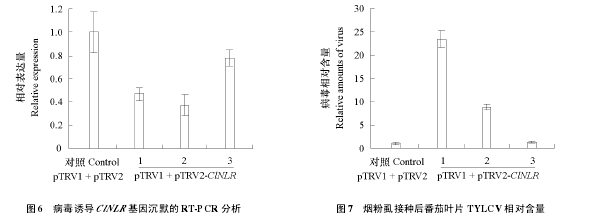
������ʱ��ȡ pTRV1 + pTRV2-ClNLR �����ķ�����Ҷ�� RNA��ͨ��ӫ�ⶨ�� PCR ��ⷢ�� 3 ����� VIGS �����ķ���ֲ�� ClNLR ����ı���ˮƽ�½������Զ��յ� 40% ~ 80%��ͼ 6������Щ pTRV1 + pTRV2-ClNLR �����ķ���ͨ���̷�ʭ���� TYLCV 7 d ��ӫ�ⶨ�� PCR ��ⷢ������ClNLR��������ʹ�������ڵ�TYLCV�������ӣ���ClNLR�������ˮƽ��������TYLCV�������ɷ��ȣ�ͼ 6��ͼ 7������ ClNLR ��������½��ķ���ֲ����ֺ�û�г��ֵ��͵�ҶƬ�ƻ��������ȷ��ѻƻ���Ҷ����֢��ͼ 5��������������� CLNLR �ij�ĬЧ��δ�ܴﵽʹ���������ȫ�����Ƶ�ˮƽ���ͱ���� ClNLR ����ά��ֲ��һ���Ŀ��ԡ�
����3�� ����
����ֲ��Բ����Ŀ��Դ�ſ�������Ϊ���������л��������ú��źŴ���;�������˲����ĸ��ƺ��ƶ����Լ��յ�������С���ӽ鵼�˿��Ի���Ļ��Culver & Padmanabhan��2007�����ֿ�TYLCV �ķ��Ӻ����������Բ������Gorovits et al.��2007���������� TYLCV �Ŀ������粢�����Ի�����������Ҫ�ġ�Ϊ�˴ﵽ���Ŀ�꣬��չ�˷�����Ӧ TYLCV ��Ⱦ��ת¼���о���������ClNLR ����Solyc05g009760.1�����ܲ���ֿ� TYLCV ��Ⱦ�ķ�����Ӧ��Chen et al.��2013�����ڴ˻����ϣ��������н�һ����¡�˸� ClNLR ����ͼ 1����ӫ�ⶨ�� PCR ֤ʵ ClNLR �����ڿ������Ѳ����ϵ�������ڸв������������Ա���仯��ͼ 2��������˵�� ClNLR �����ڿ��������п�����Ӧ TYLCV ����Ⱦ��ClNLR �����ڱ�����ҶƬ�ϵ�˲ʱ����ɲ��������Է�Ӧ��ͼ 4����VIGS ���ƿ������Ѳ��� ClNLR ����ı�������� TYLCV �ڷ���ҶƬ�еĻ��������ҷ��� ClNLR �������ˮƽ�������� TYLCV �������ɷ��ȣ�ͼ 5��ͼ 6��ͼ 7���������û�����ܲ����˷��ѿ� TYLCV �ķ�����Ӧ��ClNLR �����ڷ��ѵĸ���ҶƬ�б�����ߣ�ͼ 3����Ҳ�������俹��������ء�
����ClNLR ����ĵ����ʺ��� NBS-LRR �������������ṹ���� N ��Ϊ 1 �����͵� NB-ARC �ṹ��C �˺��� 4 �� LRR ���ؽṹ���ǵ��͵� NBS-LRR �������Ľṹ��ͼ 1����NBS ��ϸ����������ֻ���С�����䡢ϸ���Ǽܹ��ɼ�������Ӧ�ж�������Ҫ���ã�Pan et al.��2000����NB-ARC�ṹ��ֲ��������������൰���У��� GTP ��ϵ��ס�ATP �ϳ�ø β �ǻ����������������ӣ�elongation factors�����������塢R ������뵰�������ڵ����뵰��������Ϊһ����������������ã��������ʺ�����Ľ��ͨ������Ϊ�Ǹı��� R ����������źŴ���;����������Ա�������ã�Bent��1996��������������ʶ��İ�����л���λ�� LRRs�����γɶ�������������Ϊ R �����������������ϡ����뵰���뵰��Ļ�����������̼ˮ�������ϣ�����������ʶ��ԭ����Դ���źŷ��ӡ��ڶ����Ͻ� NBS-LRR ������� RPS5 �� LRR ͻ�����о��з��֣������������źŵ�ʶ�����������źŵĴ�����Wang et al.��1998�������� ClNLR �Ľṹ�������Ʋ�û������ͨ���벡ԭ��ЧӦ�ӽ��н�϶����Ʋ�ԭ��ֲ��ϸ���ж�ֲ����չ���Ӷ��ڷ��ѿ������������з������á�
������ȱ����Ӧ Avr ���������£���ʱ�� R ����Ĺ�����Ҳ������ HR ��Ӧ�����緬�ѵ� Pto ����Tang et al.��1999�������Ͻ�� RPS2 ����Tao et al.��2000�����̲ݵ� L ����Frost et al.��2004����
������Щ���ݱ����źż�����Ӧ������ȱ�ٲ�ԭ�������±��������ǿ�ҵĹ����Ի�����Ӧ�� R ���� RPS2 �� RPS5 ��ͨ�� CC ����� NB-ARC Ƭ�Σ�CC-NB-ARC���յ�������Tao et al.��2000��Juleset al.��2007�������� RPS4 �� RPP1A ���Ե����У�TIR-NB-ARC �������յ� HR��Zhang et al.��2004��Weaver et al.��2006����ClNLR ����ĵ��ײ����� TIR �� CC �ṹ���ĸ��ṹ�������� HR ��Ӧ�Լ��� R ���� HR ���߷�Ӧ���ź�ת��;�����д���һ�����о���
���������յ��Ļ����Ĭ�Ѿ���չΪ�о�ֲ������Ĭ����Ч���ߣ�Purkayastha & Dasgupta��2009����
���������õ�һ�ֲ�������Ϊ�̲ݴ��Ѳ�����Liu et al.��2002��Brigneti et al.��2004����Czosnek �ȣ�2013��ͨ���ȽϿ��з��Ѳ��ϵ� cDNA �Ŀ�� VIGS �ķ�����ɸѡ�������� 25 ���ڿ������Ѳ��������ȱ���Ļ������� 5 ������ij�Ĭʹ�����߽⣬��ָ�����Ѷ� TYLCV �Ŀ��Դ��ڶ��ι�ϵ������SlVRSLip ������ LEHT1 ��������η������ã�Sade et al.��2013������ͼ 6 ��ͼ 7 �п��Կ�����VIGS���� ClNLR �������� 1��2 ֲ����� TYLCV ���������������ӣ�ֲ�� 3 �� ClNLR �������ˮƽ�ͽӲ���Ĵ���������������������ڸû�����ֲ�� 3 ��û�б���Ч��Ĭ��ֵ��ע����ǣ��̷�ʭ���ֹ��Է���ֲ���ʹ������Ĭ�ķ���Ҳδ���ַ��ѻƻ���Ҷ����֢������������ClNLR����ij�ĬЧ�ʲ��ߣ�δ��ʹ����ı��ﱻ��ȫ���ƻ����Ƶ��в���ˮƽ��Ҳ������ClNLR����ֻ�ǿ�����Ӧ�е����η������ǿ����ؼ�����ͨ������������ԭ������û����Ƿ���й����Կ��Ը��ò��� ClNLR �����ڿ�����Ӧ�е����á�
- ��������Ƽ�
- ���ִ��ij�CsTBL1�������ӿ�¡�������2014-06-28





