���ִ��ij�CsTBL1�������ӿ�¡�������
ʱ�䣺2014-06-28 ��Դ��δ֪ ���ߣ�ɵɵ���� ����������4065��
�����������ǵ��ػ�������������Ҫ��ת¼�������ӣ�����������ʱ�ձ���ģʽ���Ծ�ȷ�ɿ��ض�λ����ı��ﲿλ�����о������ʱ�ձ���ģʽ����֤�����ܾ�����Ҫ���壨Ayoubi & van DeVen��1996�������� �ȣ�2008����DUF231 ������һ��ֲ�����е�����δ֪�ĵ��ף����൰����Ҫ�ص��ǰ������ص� DUF231�ṹ��Bouchabke-Coussa et al.��2008��Synthase��2010��Gille et al.��2011�������Ͻ��а��� 46 ��DUF231 �����Ա��ֻ�� 3 ����Ա��ESK1��AXY4/AXY4L �� TBR/TBL3������¡�������书����Ҫ�漰ֲ��淴Ӧ�Լ�ֲ�����ϸ���ڵĴ�����л��Xin��1998��Xin et al.��2007��Lugan et al.��2009��Bischoff et al.��2010�������� DUF231 ��������ܵ���Ҫ�ԣ��о������ĵ��ػ��ƾ�����Ҫ���塣�ѱ����� DUF231 ����������������֯�����Ժ�ʱ�ձ���ģʽ���� ESK1 ������Ҫ�ڴ������γɵ�ϸ����ľ�ʲ���ά������ά�ϱ��Yuan et al.��2013����TBR �������������� GUS ��������� 9 d �����Ͻ��������ҶƬë״���и߱���� 3 ~ 4 ����ֲ����ȴֻ��ҶƬά�����Ϳ��������ĸ����ϱ���� 6 �ܺ���������Bischoff et al.��2010����Ŀǰ�� DUF231 ��������ܵ��˽�ֻ������ģʽֲ�����Ͻ棬�����������л�δ��������
�����ڱ�ʵ����ǰ�ڹ������״δ�ľ��ֲ����ִ��ij��п�¡�� 1 ������ϩǿ���յ��� DUF231 ������� CsTBL1����¼�ţ�KJ094573�����û��������Ͻ� TBR �� 57%���ƶȣ������书�ܺͱ���ģʽ�в�����������ƣ�2010�������о���������Ȼ�������Ϣ���Ӱ��ִ��ij��п�¡�� CsTBL1 �������ӣ�����������Ϣѧ�ֶη����������ӵ�˳ʽ����Ԫ�����ø��������� GUS �����ںϵ�ֲ���������ת�����Ͻ棬���о���������ʱ�ձ���ģʽ������Դֲ�D�ص���Ӧ��Ϊ�����˽� CsTBL1 ��ֲ��������еı���ģʽ�ṩ�������ݡ�
����1�� �����뷽��
����1.1 �������
����������2012—2013�����й�ũҵ��ѧԺ�̽��о�����Դ�ҽ��С����ִ��ij�[Citrus sinensi��sL.��‘Olinda’]ҶƬ�����й�ũҵ��ѧԺ�̽��о�����Դ�ԡ�pEASY-T1Vector ���Ա���ȫʽ�����\������˾���˾� DH5α��ũ�˾� LBA4404���������� pBI121 �ɱ�ʵ���ұ��档
��������ת���ĸ��ױ��ǣ�Columbia��Co1��Ұ�������Ͻ��� 22 �棬16 h �ڰ�/8 h ����������Ӫ������ 4 �ܣ�Ȼ���� 22 �棬8 h �ڰ�/16 h ������������ֳ������ʢ���ڡ�
����1.2 DNA �� RNA ����ȡ�� cDNA �ĺϳ�
��������ҶƬ�����Ͻ�ֲ��� DNA С����ȡ���������˾ Plant Genomic DNAKit ˵���������ʹ�� QIANGEN ��˾ DNeasy Plant Maxi Kit ��ȡת�������Ͻ������ DNA ���� Southern �ӽ�����ȡ���̰�����˵������С�ʹ�������˾ RNAprep pure Kit �Լ�����ȡ���Ͻ�ֲ���� RNA��ʹ��TaKaRa ��˾ PrimeScriptTMRT reagent Kit ��ת¼�Լ��кϳ� cDNA��
����1.3 ���������п�¡�����������幹��
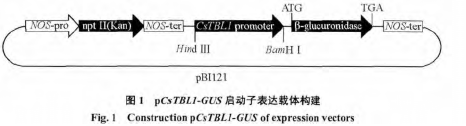
������ CsTBL1 cDNA ȫ������Ȼ��������н��� Blastn �ȶԻ�� CsTBL1 ���������Ӳο����С����òο�����������P1/P2���� 1�������ִ��ijȻ�����DNA Ϊģ�壬�� PCR ������õIJ������ӵ� pEASY-T1�����ϣ�����������˾������������ο��������� DNAStar �������бȶԷ��������� PLACE ���ݿ���������������еĵ���Ԫ����Lescot et al.��2002����������ȷ��ת�������� Hind ��� BamH��˫ø���滻 35S �����ӣ����ӵ����� GUS ����� pBI121 �����ϣ���������pCsTBL1-GUS��ͼ 1��������ø�кͲ��������������� PF/PR���� 1�������ȷ�������������ö��ڷ�ת��ũ�˾� LBA4404��
����
����1.4 CsTBL1 ����������ת�����Ͻ�
��������ũ�˾��鵼�Ľ��������������� pCsTBL1-GUS ת�����Ͻ棨Zhang et al.��2006��������õ� T1���Ͻ��������������ں��� 50 mg · L-1Kan������ù�أ��� MS ��������������������ѡ����ɫ�����磬�����Ե�Ӫ�����С���ȡ T1��������� DNA���� Kan ����λ������������� NPT��NPT��F/NPT��R�������С�� 589 bp���� 1�����ת����ֲ�ꡣ�� NPT������ PCR ������������̽�룬�� T3��ת�������Ͻ濽�������� Southern �ӽ�������
�������巽������ Roche ��˾������“DIG High Primer DNALabeling and Detection Starter Kit��”�Լ���˵������С�
����
����1.5 GUS ��֯��ѧȾɫ
�������� Ni �ȣ�2008���� GUS ��֯��ѧȾɫ������ȾҺ��ȡ��ͬ����ʱ��ε����Ͻ���֯���� GUSȾҺ�� 37 ���ҹ���� 75%�ľƾ���ɫ����ȫ��Ȼ���������¹۲����ա�
����1.6 ��Դ���ش�����ת�������Ͻ� GUS ����ı������
����������20 d��T3��ת�������Ͻ���100 mol · L-1ABA��20 mol · L-1ACC��200 mol · L-1MeJA��20 mol · L-1SA ���� 12 h ���ռ���Ʒ–80 �汣�档ʹ�� qRT-PCR ���� GUS ����ı�������GUS��GUS-F/GUS-R����������Ͻ��ڲ����� Tubulin��Tubulin-F �� Tubulin-R����Oate-Sánchez et al.��2007������ 1������ʵʱ PCR ʹ�� Promega GoTaqqPCR Master Mix���� Bio-Rad CFX96TMӫ�ⶨ�� PCR �ǣ�Bio-RAD��USA���Ͻ��С�PCR ��Ӧ��ϵ��20 L������ 10 L Master Mix��2×����0.2 L�������10 mol · L-1����0.2 L �������10 mol · L-1������ cDNA1 L��PCR ��Ӧ����Ϊ 95 ����� 2 min �����²���ѭ�� 40 �Σ�95 �� 10 s��60 �� 60 s�������ܽ����ߣ��Լ�������Ƿ���á����� 3 ������ѧ�ظ��� 3 �������ظ������ݷ������չ�ʽ��2-Ct��Ct = CtĿ�����–Ct�ڲλ����㡣�趨δ�������ϵ����ݣ����գ�Ϊ 1������������Խ��бȽϡ�ʹ�� Spass19.0 �����������������Է�����
����2�� ��������
����2.1 ���п�¡�������ͱ�������Ĺ���
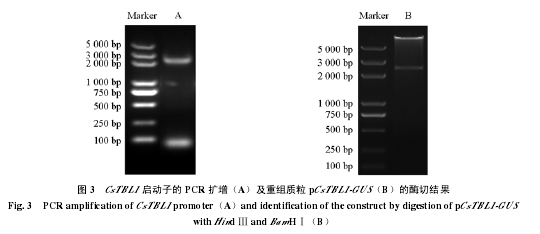
������ CsTBL1 cDNA ȫ����������Ȼ��������н��� Blastn�ȶԣ������ CsTBL1 �Ļ������������Ӳο����У����ݴ����������Ӱ��ִ��ijȵĻ����� DNA�п�¡���û���������ӡ�������������õ������ӳ�Ϊ 2 361 bp��ͼ 2������ο����е����ƶȴﵽ 98%����������վ PLACE ���ݿ����CsTBL1 �������������У�������� 2���ɼ��������Ӻ������澳��ֲ�D��Ӧ����ص�˳ʽ����Ԫ�� ABRE��ASF��GARE��GT1 �� MYB �ȡ�
����

�����������Ӿ�˫ø�д�TA��¡��ж�أ����Ҿ����Ӳ����ɹ��滻��pB1121�����ϵ�35S�����ӣ���ͼ 3 ��ʾ������õ� pCsTBL1-GUS �������徭�������˫ø�У�֤ʵ������ȷ��
����
����2.2 ת�������Ͻ�Ļ�ü���֤
����ͨ�� Kan ����ɸѡ������� 8 �꿹��ת��ֲ�ꡣ�� NPT������������õ� T1��ת����ֲ�꣬�������п���ֲ�����ת�������ԣ�ͼ 4����˵������ɸѡ��Ч���� 6 �� T2��ת������ϵ�Ŀ�����ǿ��������Ϊ 3�U1��������Ϊ�ǵ��������롣��ת������ϵ��� T3���� Southern �ӽ���֤��ͼ 5������֤����� 2��3��5��6��7 �� 8 ����ϵȷʵΪ������ T-DNA ���롣
����
����2.3 ��ͬʱ��ת CsTBL1 ���Ͻ�ֲ��� GUS ��֯��ѧȾɫ
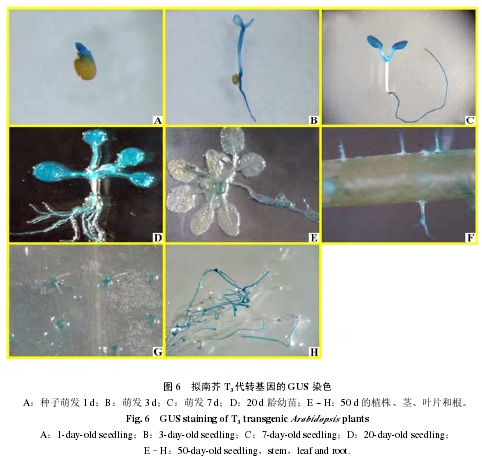
����ѡȡ�ȷ� 1��3��7��20 �� 50 d �� T3������������ת������ϵ���� GUS Ⱦɫ��������������Ӹո��ȷ� 1 �� 3 d ʱ GUS ��������ߣ�ͼ 6��A��B�����ȷ� 7 �� 20 d ���������Ҷ����Ҷ����GUS ����Ҳ��ǿ��ͼ 6��C��D�������������� 50 d ��ת�������Ͻ��� GUS �����½���ֻ��ҶƬ�;���ë״���Լ������ĸ����б��ͼ 6��E ~ H����˵�� CsTBL1 �����Ͻ��еı������ʱ�ղ��죬��Ҫ�����������Լ����ڷ�������֯�б��
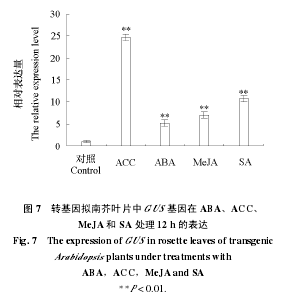
����2.4 ת CsTBL1 ���������Ͻ��� GUS ������ ABA��ACC��MeJA �� SA �����µı����ò�ֲͬ�D�ض�����20 d��ת�������Ͻ����紦�� 12 h�������ҶƬ�� GUS ��ת¼ˮƽ�������ͼ 7����ʾ����ϩ�ϳ�ǰ�� ACC��1–����������–1–���ᣩ����ǿ���յ�GUS �ı������֮ǰ��оƬ������ʾ��CsTBL1 ��������ϩǿ���յ��Ľ���������ƣ�2010����һ�¡���Դ ABA�������ᣩ��MeJA���������ᣩ�� SA��ˮ���ᣩҲ�����յ� GUS����ı��
����3������
�������ѱ�������������Ҫ��Ϊ 3 ���ࣺ����͡��յ��ͺ���֯�����ͣ�Carninci et al.��2006��Li et al.��2006������֯����������������������һ����Ҫ�����ͣ���ṹ���ͱȽϸ��ӣ����������ӿ���Ŀ�Ļ������ض���֯�����ٱ���������ػ����������ֳ����ض��Ĵ�л�����ʱ����ص��ص㣨���� �ȣ�2007��÷����2008�����壬2013�������о���ʾ��CsTBL1 ���������� GUS ��������� 1 ~ 3 d ������������� 7 ~ 20 d �������ҶƬ����ǿ�ұ������ֲ��ķ��� GUS ���Խ��ͣ�50 d ��������ֻ��ҶƬ�;���ë״���Լ������м� GUS ���ԡ����������Ͻ�ͬԴ���� TBR �����ӵı���ģʽ���ƣ���������������ҶƬ�;���ë״���д������������ֲ����ֻ�ڿ���������λ���Bischoff et al.��2010������˵�� CsTBL1 �������ʱ�պ���֯�����ԣ���ֲ�����ڷ���������ء�
������֯�����������Ӳ�������һ�������ӵĽṹ���������������п���������֯����ĵ���λ�㣨Guilfoyle��1997��÷����2008���������ѱ����������ӷ�������֯�����������ӵ�˳ʽ����Ԫ����ɱ����ԣ��� GTGA �� TGTGG �����ǻ������Ա���Ԫ������ӯӯ �ȣ�2013����B-box Ԫ�������������������и߶ȱ��أ�Ezcurra et al.��1999����CsTBL1 ����������˳ʽԪ�������������ж����ë״�����������ص�˳ʽ����Ԫ�� MYB��Wang et al.��2002��Ni et al.��2008��Shangguan et al.��2008�����ڸ��ٵ���Ҷ�����ҡ������������ʵ��Ҳ����ë����ë�ķ����Ƿ�Ҳ�� CsTBL1 �IJ��������һ�����о���
����CsTBL1 ������������Ϣѧ������ʾ�京������澳Ӧ���������Ԫ������ ASF��ABRE��GT1MYC �� GARE �ȣ��Ʋ�������ӿ�����Ӧ����������ӡ�������/��ϩ;����ˮ��������;����ֲ��������Ҫ�ķ���Ӧ���ź�;����Pieterse et al.��2001��Horváth et al.��2007��Loake & Grant��2007����������������Դ ACC��MeJA �� SA �Ĵ��������������� GUS �ı�������֤ʵ���������۵���ȷ�ԣ�ͬʱҲ˵���� CsTBL1 �����������ַ����ź�;��������ڡ�ֵ��ע����ǣ�ǰ���о����������֤�� CsTBL1 ����ϩǿ���յ�������ϩ�ź�;���е����λ����� CsTBL1 ������������ϢԤ��ȴû�з�����ϩ��صĵ���Ԫ�����˽����ʾ�� CsTBL1 �������д�������ϩ���ص�δ֪Ԫ�����д��ڽ�һ���ھ�
- ��������Ƽ�





