
哮喘是一种免疫调节异常的呼吸系统疾病,其发病机制复杂.免疫反应异常是哮喘发生的重要环节,在哮喘中发挥关键作用[1].本课题前期构建了热休克蛋白 70 ( heat shock protein70,HSP70) /CD80 DNA 疫苗,以该重组融合蛋白作为免疫原,刺激机体产生免疫应答.并观察 HSP70/CD80DNA 疫苗对小鼠急慢性哮喘的预防和治疗作用,发现其机制可能是从非特异性免疫方面恢复辅助性 T细胞 1 ( T helper cell 1,Th1) /辅助性 T 细胞 2(T helper cell 2,Th2)平衡[2-3].随着人们对 CD4 +T 细胞认识的逐渐深入,发现 CD4 + CD25 + 调节性T 细胞( regulatory T cell,Treg) 和辅助性 T 细胞 17(T helper cell 17,Th17)也在哮喘的发生中具有重要作用.HSP70/CD80 DNA 疫苗在治疗哮喘的过程中是否也调节 Treg /Th17 的平衡,目前尚不清楚.
本研究拟探讨 HSP70/CD80 DNA 疫苗治疗小鼠急性哮喘的作用及其对 Th1/Th2/Treg /Th17 的影响,旨在为该疫苗的临床应用提供实验基础.
1 材料与方法
1. 1 材料 健康雌性、SPF 级近交系 BALB / c 小鼠购于第三军医大学; HSP70/CD80 DNA 质粒由泸州医学院附属医院感染与免疫实验室构建[4];End-ofree 质粒大量提取试剂盒购于德国 QIAGE 公司;离心柱式总 RNA 提取试剂盒购于天根生化科技(北京)有限公司;iScriptTMcDNA Synthesis Kit 购自美国 Bio-Rad 公司;TaKaRa SYBR Premix Ex TaqTMII 购自宝生物工程( 大连) 有限公司;Real-time PCR引物由宝生物工程(大连)有限公司合成.
1. 2 方法1. 2. 1 疫苗制备 pVAX1 ( +) 质粒和 HSP70 /CD80 DNA 质粒的制备按照试剂盒说明书进行.测定纯度及浓度后,用生理盐水调整浓度到1 mg/mL备用,-20 ℃存储.
1. 2. 2 动物模型制备 BALB / c 小鼠随机分为 4组:空白对照组(空白组),哮喘模型组(模型组),pVAX1(+) 空载体对照组( 空载组) 和 HSP70 / CD80疫苗治疗组(治疗组),每组 10 只.模型制备过程中,空载组死亡 1 只小鼠.实验开始第 0 和 14 天,新鲜配制含 10 μg 鸡卵清蛋白(ovalbumin,OVA)的铝佐剂混悬物 200 μL,分别腹腔注射模型组、空载组和治疗组小鼠,而空白组小鼠注射等剂量生理盐水.第 17 和 25 天,治疗组小鼠股四头肌处肌内注射 100 μg HSP70/CD80 DNA 疫苗,空载组注射100 μg pVAX1,空白组和模型组注射等体积生理盐水.第 28 天,对模型组、空载组和治疗组小鼠以1% OVA 行雾化吸入激发,连续 7 次,30 min / 次.空白组生理盐水雾化.
1. 2. 3 气道反应性检测 最后一次 OVA 雾化 24 h内,吸入乙酰甲胆碱(methacholine,Mch)后检测小鼠肺功能,全身体积描记仪记录 Penh,按以下公式计算出%基线 Penh 值:100 × (检测 Penh 值 - 基线Penh 值) / 基线 Penh 值,各组进行比较.
1. 2. 4 ELISA 法检测血清 IgE 取小鼠眼球血,4 000 r / min离心 10 min,收集血清,- 80 ℃ 保存.
1. 2. 5 观察肺组织病理学改变 切取肺组织固定于4%多聚甲醛12 h.制备切片,进行 HE 和AB-PAS染色.镜下观察炎细胞浸润、杯状细胞增生分泌情况,参照文献对炎细胞浸润和杯状细胞进行评分[5-6].
1. 2. 6 ELISA 法检测气管肺泡灌洗液( bronchoal-veolar lavage fluid,BALF)中细胞因子含量 注入400 μL 生理盐水到主支气管,反复灌洗 3 次,吸出BALF,2 000 r / min 离心 10 min,收集上清,检测 γ-干扰素(interferon-γ,IFN-γ)、白细胞介素-4(inter-leukin-4,IL 4)、转 化 生 长 因 子 β ( ransforminggrow th factor beta,TGF-β)和白细胞介素-17(inter-leukin-17,IL 17)含量.
1. 2. 7 检测肺组织中转录因子 mRNA 水平 按试剂盒说明书进行肺组织总 RNA 提取、逆转录和Real-time PCR 反应.检测各样本 T-bet、GATA 连接蛋白 3(GATA binding protein 3,GATA3)、叉头蛋白 3(forkheah box protein3,Foxp3)和维甲酸受体相关孤儿受体 ( retinoid related orphan receptorgamma t,RORγt) Ct 值.根据 Ct 值,用软件 ABIStepOne 进行分析,按公式 2- △△CT计算各样本相对表达量.
1. 3 统计学处理 采用 SPSS 17. 0 软件.先用Shapiro-Wilk 行正态分布检验,符合正态分布即可用方差分析行多组间比较,用 LSD 检验行多组间两两比较,数据以x± s 表示.如不符合用 Kruskal-Wallis H 检验行多组间比较,用 Mann-Whitney U 检验行多组间两两比较,数据以 M(P25,P75) 表示.P < 0. 05 为差异有统计学意义.
2 结 果
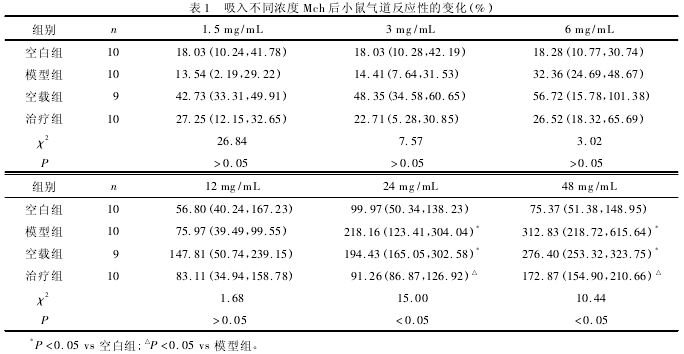
2. 1 气道反应性的改变 见表 1.Mch 吸入浓度达到 24 mg /mL 后,与空白组小鼠相比,模型组和空载组小鼠气道反应性升高 (P <0. 05);与模型组小鼠相比,治疗组小鼠气道反应性降低(P <0. 05).
2. 2 IgE 水平的变化 模型组和空载组的血清 IgE浓度[(7.45 ±0.81) pg/mL,(7. 08 ±0. 74) pg/mL]高于空白组[(5. 36 ±0. 29) pg /mL],P <0. 05;治疗组的血清 IgE 浓度[(5. 91 ±0. 52) pg /mL]低于模型组和空载组(P <0. 05).
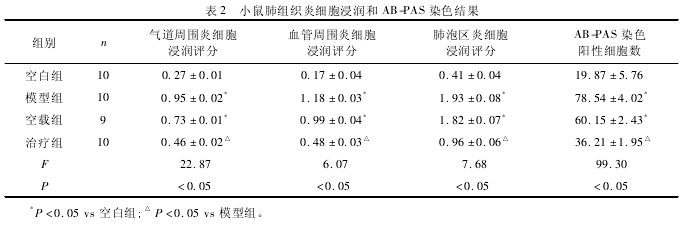
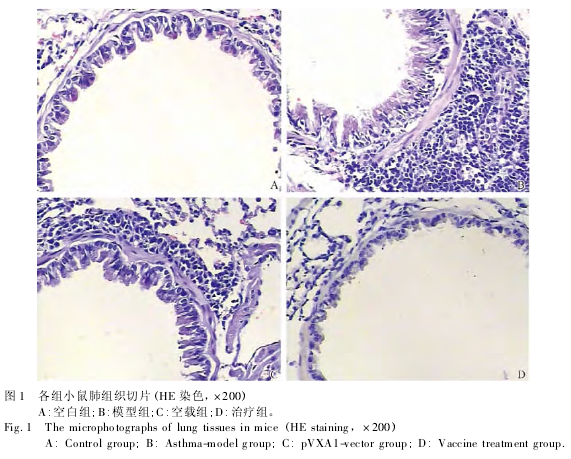
2. 3 肺组织炎细胞浸润和黏液分泌情况 见表 2、图 1.与空白组相比,模型组和空载组小鼠肺组织出现大量炎细胞浸润(P <0. 05),以单核细胞和嗜酸粒细胞为主;与模型组和空载组相比,治疗组小鼠的气道、血管周围和肺泡区炎症反应减轻 (P <0. 05) .AB-PAS 染色后进行杯状细胞评分,结果表明,模型组和空载组小鼠的支气管上皮中阳性杯状细胞增多(P <0. 05);与模型组相比,空白组和治疗组小鼠杯状细胞增生不明显(P <0. 05).
2. 4 细胞因子的改变 见表 3.计算 BALF 上清液中细胞因子 IFN-γ/IL-4 和 TGF-β/IL-17 比值,结果表明,模型组的比值低于空白组和治疗组比值(P <0. 05).
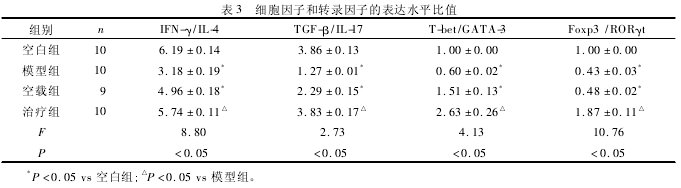
2. 5 转录因子表达水平 见表 3.计算 T-bet / GA-TA-3 比值和 Foxp3 / RORγt 比值,结果表明,模型组比值低于空白组和治疗组比值(P <0. 05).
3 讨 论
哮喘以气道高反应性、气道炎症和气道重建为主要特征.本研究用 OVA 致敏激发小鼠,末次雾化激发后小鼠出现上述典型症状,说明模型成功建立.HSP70 是一类以分子伴侣形式维持细胞稳态的高度保守的蛋白.HSP70 可以通过干扰细胞内炎症信号转导通路而抑制炎症反应[7].在哮喘的炎症反应中,HSP70 与树突细胞、单核细胞和髓系衍生抑制性细胞表面受体相结合,从而使这些细胞免疫应答活化.HSP70 还可增强 Treg 细胞因子IL-10和 TGF-β 的分泌[8].CD80 是免疫球蛋白超家族成员,常表达于 DCs、B 细胞、巨噬细胞等细胞表面.研究表明,CD80 通过与 T 细胞表面分子CD28 和 / 或 CTLA-4 作用,为 Treg[9]、Th17[10]等 T淋巴 细 胞 活 化 提 供 共 刺 激 信 号.HSP70/CD80DNA 疫苗对急性哮喘小鼠进行干预后,发现哮喘气道症状有所减轻,提示该疫苗可能对其有治疗作用.
Th1 / Th2 失衡是哮喘发病的核心.Th2 活化过程中,转录因子 GATA-3 和 T-bet 发挥重要作用[11].T-bet 是调节 Th1 分化的关键转录因子,它能抑制IL-4 的合成[12]和 Th2 的活化[13].哮喘时 Th2 增强,Th1 抑制,Th1/Th2 平衡向 Th2 发生偏移.本研究用 HSP70/CD80 DNA 疫苗治疗哮喘小鼠后,发现治疗组 IFN-γ/IL-4 比值和 T-bet/GATA-3 比值相较模型组升高,IFN-γ 和 T-bet 也升高,说明该 DNA疫苗上调 IFN-γ/IL-4、T-bet/GATA-3,可能通过改变 Th1/Th2 失衡状态,从而发挥对哮喘的治疗作用.
Treg 细胞能够抑制哮喘反应的发生.Treg 细胞分泌 TGF-β、IL-10 等细胞因子,抑制嗜酸性粒细胞、嗜碱性粒细胞和肥大细胞的活化,减轻 Th2 细胞因子 IL-9 等的分泌,从而抑制气道炎症反应.
Th17 细胞能够促进哮喘反应的发生.Th17 细胞合成、分泌细胞因子 IL-17,IL-17 具有活化支气管上皮细胞的作用,使其分泌多种促炎因子,从而加重气道炎症反应[14-15].Treg 和 Th17 细胞相互制约,维持平衡.转录因子 RORγt 和 STAT3 调控 Th17 细胞的活化.Treg 分化由关键转录因子 Foxp3 调节和信号转导通路 TGF-β/Smad 调控.本研究结果显示,模型组 TGF-β/IL-17 比值和 Foxp3/RORγt 比值降低,同时 Foxp3 降低、RORγt 升高,提示免疫平衡向Th17 发生偏移.通过疫苗治疗后,降低的 TGF-β /IL-17 比值和 Foxp3 / RORγt 比值回升,TGF-β 和Foxp3 也升高,说明 HSP70 / CD80 DNA 疫苗可能通过改变 Treg /Th17 平衡,而在哮喘的治疗中发挥作用.
Th1 / Th2 / Treg / Th17 细胞间并非是静止不变的,而是保持着一种动态平衡关系.与以往的研究结果一致,本研究发现 Th2、Th17 的促炎反应与Th1、Treg 的抑制炎症之间存在相互协同、相互抑制的作用.HSP70/CD80 DNA 疫苗除了通过 IFN-γ/IL-4 和 T-bet / GATA-3 比值的上调,以恢复 Th1 /Th2 平 衡 外,还 能 上 调 TGF-β / IL-17 和 Foxp3 /RORγ 比值,从而恢复 Treg / Th17 平衡,最终缓解哮喘的各种气道症状,对其产生治疗作用.
参考文献:
[1] Pelaia G,Vatrella A,Maselli R. The potential of biolog-ics for the treatment of asthma[J]. Nat Rev Drug Dis-cov,2012,11(12) : 958-972.
[2] 成争艳,史小玲,何光彤,等. 热休克蛋白 70 / CD80 嵌合疫苗对支气管哮喘小鼠的影响[J]. 中华结核和呼吸杂志,2009,32(9):716-717.
[3]李燕,郭宇,史小玲,等. HSP70/CD80 DNA 疫苗通过调节趋化因子及受体对哮喘小鼠气道炎症的作用[J]. 中华微生物学与免疫学杂志,2013,33(2): 123-128.LI Yan,GUO Yu,SHI Xiaoling,et al. The effect ofHSP70 / CD80 DNA vaccine on airw ay inflammation byregulating the chemokine and the receptor of chemokine inasthmatic mice[J]. Chin J Microbiol Immunol,2013,33(2):123-128.
[4]李晖,史小玲,钟森,等. hsp70/CD80 嵌合 DNA 真核表达质粒的构建及鉴定[J]. 中华结核和呼吸杂志,2002,25(4): 244-245.
[5]Tanaka H,Masuda T,Tokuoka S,et al. The effect of al-lergen-induced airw ay inflammation on airw ay remodelingin a murine model of allergic asthma[J]. Inflamm Res,2001,50(12): 616-624.
[6] McMillan S J,Xanthou G,Lloyd C M . Manipulation ofallergen-induced airw ay remodeling by treatment w ith an-ti-TGF-beta antibody: effect on the Smad signaling path-w ay[J]. J Immunol,2005,174(9): 5774-5780.
[7] Wakashin H,Hirose K,Maezaw a Y,et al. IL-23 andTh17 cells enhance Th2-cell-mediated eosinophilic airw ayinflammation in mice[J]. Am J Respir Crit Care Med,2008,178(10): 1023-1032.
[8] Wachstein J,Tischer S,Figueiredo C,et al. HSP70 en-hances immunosuppressive function of CD4(+) CD25 (+)FoxP3(+) T regulatory cells and cytotoxicity in CD4(+)CD25(-) T cells[J]. PLoS One,2012,7(12): 51747-51777.
[9]Barnes M J,Griseri T,Johnson A M,et al. CTLA-4promotes Foxp3 induction and regulatory T cell accumula-tion in the intestinal lamina propria[J]. Mucosal Immu-nol,2013,6(2): 324-334.
[10]Bouguermouh S,Fortin G,Baba N,et al. CD28 co-stimulation dow n regulates Th17 development[J]. PLoSOne,2009,4(3): 5087-5117.
[11] Maier E,Duschl A,Horejs-Hoeck J. STAT6-dependentand -independent mechanisms in Th2 polarization[J].Eur J Immunol,2012,42(11): 2827-2833.
[12]Lohoff M,Mak T W. Roles of interferon-regulatory fac-tors in T-helper-cell differentiation[J]. Nat Rev Immu-nol,2005,5(2): 125-135.
[13] Oestreich K J,Weinmann A S. T-bet employs diverseregulatory mechanisms to repress transcription [J].Trends Immunol,2012,33 (2): 78-83.
[14] Oboki K,Ohno T,Saito H,et al. Th17 and allergy[J]. Allergol Int,2008,57(2): 121-134.
[15]李洧,叶茜,高顺翠,等. 白细胞介素 17 抗体对哮喘小鼠中性粒细胞的影响[J]. 山东大学学报:医学版,2009,47(3) : 43-47.LI Wei,YE Qian,GAO Shuncui,et al. Effects of IL-17 neutralization antibody on polymorph-nuclear of asth-ma in mice[J]. Journal of Shandong University: HealthSciences,2009,47(3): 43-47.





