����ժҪ������ֲ�����ֶ�ȫ����̬ϵͳ����������в,��������ֲ��Ĺ��������ȡ����һ����չ,��Ҫ�ָ���������̬ϵͳ����ɹܿ�Ŀ���Դ������⡣���Ķ�����ֲ��ܿس�Ч����Լ���ؽ���������,��������ֲ��ܿع����д�����������:(1)Ԥ��Ϊ���Ĺ���ԭ����ȫ�������ֹ���;(2)����ֲ���������ȱ������֧�֡���̬�������嵥һ����ͷ������ȱ��;(3)����ֲ�����Ŀ����������Ŀ�꼰��̬��������Ŀ��ì��;(4)����ֲ����ƴ�ʩ�ɱ��߰�,���ܻ�����������,������������̬ЧӦ;(5)Ŀ������ֲ����ƺ���ܲ�����������ЧӦ�������̷����֡�����,ȫ��仯�ڼ���ֲ�����ֵ�ͬʱ��ı��������,��һ�������˹ܿ��Ѷȡ��ݴ�,������Ϊ,δ��Ӧ��ǿ��ʱ�ճ߶�������ֲ����̬ЧӦ�����������ָ����ܿؾ��ߵȷ�����о�,����������ֲ��ܿس�Ч,�ָ�ֲ��������̬ϵͳ��
�����ؼ��ʣ���������; �����ָ�; ȫ��仯;
����Restraining factors on the effectiveness of invasive plants management and control
����XU Guang-yao LI Hong-yuan MO Xun-qiang MENG Wei-qing
����College of Environmental Science and Engineering,Nankai University School of Geographic and Environmental Sciences,Tianjin Normal University
����Abstract��Biological invasion is one of the major causes of global biodiversity loss. Although some progress has been made in its management and control,there are still problems in restoring the invaded ecosystems and completing the management objectives. We reviewed the restraining factors on invasive plant management and control. The following problems are found in the management of invasive plants:( 1) the precautionary principle is not fully applicable to invasion managements;( 2) invasion management is facing the problems of lacking public participation,single ecological responsibility subject,and multiple management;( 3) target conflicts exist in management of invasion management and social management and ecological environmental protection;( 4) measures on invasive plants control are costly,susceptible to environmental factors,and common to produce side effects;( 5) legacy effects and secondary invasion emerged after target plants are controlled. In addition,global change may exacerbate plant invasion and change the concept of human beings,which will further increase the difficulty of control. Hence,to enhance the effectiveness of management and control efforts and restore invaded ecosystems,it is necessary to strengthen the research on ecological effects of invasive species,invaded habitat restoration,and management decision at large spatial and temporal scales.
�����������ֱ���Ϊ�ǽ�������Ϣ���ƻ�������������Լ��ٵĵڶ���ԭ��,ֲ������̬ϵͳ������Ҫ��������,����ֲ������������̬ϵͳ��ռ�����Ƶ�λ,�����ı���̬ϵͳ�����й���(����ҫ��,2018b)�����ܲ�ͬ����ֲ���������̬ϵͳӰ��ij̶Ⱥͷ�����������,��ֲ�������ձ齵���˱���ֲ��ķ��(-43.5%)��������(-50.7%)���ʺ϶�(-41.7%),����ֲ�����̬ϵͳ��Ӱ�컹������ʳ������Ӫ��������,���ͱ��ض�����ʺ϶�(-16.5%)���ḻ��(-17.5%)(Eviner et al.,2012)����ȫ���ƶ���,����ֲ���������������,Kleunen��(2015)��ȫ����������ֲ�����������ͳ��,������13168�ֹ黯ά��ֲ��,���ع�������10%���Ϊ����ֲ�
������ǰ,��ֲ�����ֵĶ��������֡����ֻ��Ƽ���̬ѧЧӦ�����ֿ���������ȷ�����о���ȡ����Ҫ��չ(����ͤ��,2012),������ֲ����о����ն�Ҫ����ʵ��Ӧ�á�ֲ���������º����,����ֲ����϶��ǻ�������ɵĸ���Ӱ��,���ܶ��������DZ��������ֲ��,�ڳɹ�����ǰ���ڽϳ���DZ����,��������ֲ������ʶ�����ڿ����쳣����(Simberloff,2014),ʹ������ֲ��ķ��س�Ϊ������̬ϵͳ�ָ�������ֶΡ�����ֲ��Ŀ��ƾ�����Դ��ũҵϵͳ���Ӳݿ���,���ڸ����ӵĻ���(��������Ȼ�����ͳ��л���)��,���ա����εȻ��������Լ��Ļ������õ��������������Լ���Ƴ�Ч(Kettenring et al.,2011)������ȫ������仯���������÷�ʽ�ı��ԭ��,����ֲ�����������Ӱ���ͬʱ,�������ṩ��������̬������(Tassin et al.,2015),���������Ŀ���ͻ�������������ͬ�������ŵĹ������߳�ͻ(Mason et al.,2017)�����۹�ģ�ϵ�����ֲ��ܿ���Ҫ����IJ���Ͷ��,�����Ƹ����á�����仯������ЧӦ���̷����ֵ�����ע�������Ա�֤���ֿ���Ч��,���һ�������˹ܿ��Ѷȡ���ȫ������������ȵ����֮һ���ϷǸ����ն���(Cape Floristic Region,CFR)Ϊ��,ֲ������ʹCFR�ر���������30%����,��ǧ�ֱ���ֲ���ܵ���в,Ϊ�������������,�Ϸ�����������Ϊ�ڳ���20�������ֲ����ƹ���,Ȼ��,����С��Ⱥ������Ѽ����Ƴɱ�̫��,����ֲ����δ����(van Wilgen et al.,2016)������������Ӱ��ֲ�����ֹܿس�Ч������,������ز������Է�����Ȩ��,Ϊ�������ֲ��ܿس�Ч���ٽ�ֲ������������̬�ָ��ṩ�ο���
����1 ����ֲ����������е�����
����1.1 Ԥ��Ϊ���Ĺ���ԭ������ʵ��
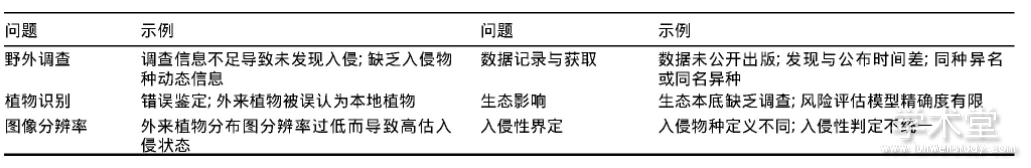
����ֲ������Ƿ�ֹ����ֲ�����ֵĵ�һ������,�����������Ǽ��ߵĻ���������̬ѧ�Ƕȿ�,����ʹ�ð������ƶ�(ֻ�������Էdz��͵����ֲ�������),��������ȫ��ֹ����ֲ������,�����ھ�����ᷢչ����Ҫ,���й���ʹ�õĶ��Ǻ������ƶ�(������������)����ʹ����,�����϶�����ͳһ����̬��Ϣ���²���ʱ��ԭ��������ֲ����������Ѹ�ٸ��¡�
����1.1.1 ����ֲ���϶�����ͳһ
���������������������Ƿ�����������Ԥ������������ֲ��Ļ���,Ȼ��,��ͬѧ�߶����ָ������ʶ��ͬ,��������ֲ����϶�������ȷ(Courchamp et al.,2017),������γ�ͳһ������,��Ϊ����ֲ���������ϰ���ֲ��������ͨ����������5��ά�����ж�:�ֲ����(local abundance)�������ֲ���(geographic range size)�������ֲ���(environmental range size)����ɢ����(spread rate)������ЧӦ��Ӱ��(negative effect or impact)(Catford et al.,2016),�����ж����ݵ�ȱ�����²�ͬ�о���Ա���ܽ�ͬһ����������ͬ���������ֹ����ڲ�ͬ������״̬������,��������ʹֲ�����ֵĽ綨���Ӹ���:����ֲ�������ʱ����ֵ��ν綨,����������ǧ��ǰ�����ֲ���㲻������ֲ��?����ֲ��Ӧ�û��ں��ֱ߽�綨,�ǵ����߽�(γ�ȴ�)�������α߽�(����)?���������α߽�,�Ƿ�Ӧ�ð���ȫ��仯���µġ������ġ���Χ��ɢ�ı���ֲ��(Nackley et al.,2017)?��ɶ�����̬�����û������ʧ������ֲ���������ֲ��?
����1.1.2 ����ֲ�ﶯ̬��Ϣ���²���ʱ
���������������º����,ֻ������ֲ����ɺܴ�ĸ���Ӱ��ʱ,�����϶���Ϊ����ֲ��,�������ֲ�������ĸ��±�Ȼ�����ͺ���(��1)������Ⱥ�䶯̬��,��Ҫ���ڵ����Ա���ȷ������ֲ����Ϣ,������ֲ�������,ȫ����տ�(Compositae)���̱���(Poaceae)�Ͷ���(Leguminosae)ֲ����,�黯���־ͽ�4000��(Py2ek et al.,2017),�����о���Դ�����ܷ��䵽ÿ�����֡�ÿ������������ֲ�ﱻ�����,������ɢ������ɽ�صȽ�ͨ����ĵ���,�����ڴ���С��Ⱥ��,����ֲ���ʵ�ʷֲ���̬���Ѿ����(Hyndman et al.,2015)������,ijЩ����ֲ���뱾��ͬ���ֻ��Ե�ּ�������,��ʹרҵ��ֲ��ѧ��Ҳ�����ж�,���������ϵ��ѷ���ֲ�����ѧ�IJ�ȷ��ʹ�������������(Hirsch et al.,2017)��
������1 �����������������д��ڵ�����
����1.2 ����ֲ������ƶȴ���ȱ��
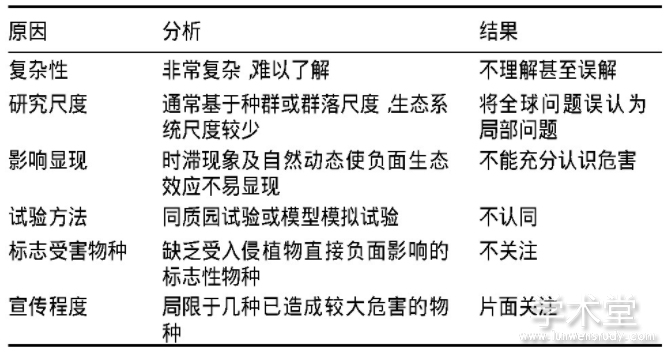
����������������̬�������ֹ��̵ĸ�����,���ں��������֧�����ֹ���������ֲ�・��ʽ��̬Ӱ�켰��;����ɢ��ʽ,ʹ��̬�������������ȷ��,����Ȩ��Ļ��ҽ�һ������������ֲ��Ĺ�����Ч��
����1.2.1 ���ڲ����ƶ�����ʵ
�������������ͰĴ����ǵ�����������,�������ں������ӻ����ֲ�����֡�û����̬ѧ�����Ĺ��ڻ����Ϊ����ֲ�������̬ϵͳ�����������,������ij��ֲ���Ա������������ȴ�ǻ�����(��2)(Courchamp et al.,2017)���о��Ϳ�������ֲ��ĸ���ԭ���Ȼ���������̬�;�����ʧ,��Ҫ����̬ϵͳ����Ȼ�仯����������ֲ��Ľ���ʽӰ��dz�����,��ʵ��,��ʹ�Ƕ���������ǿ��ֲ����ɵ����ֱ�۹��Ƶľ���Ӱ��,����Ҳ֪֮���١������������������(��ȫ���ů�뱱����,���Ȳ����뾨��),�������о���������ֲ�����ϧ����Σ���������Ӱ��,�����ɽ��������ۼ����ڶ�����ֲ��Ĺ�ע(Touza et al.,2014)��
������2 ����ֲ�����ȱ�����ڲ����ԭ�����
����1.2.2 ��̬������������ȷ��
����ֲ�����ֵ�������������ֻ��Ҳֻ���ǹ���(����),90%���ϵ�����ֲ������Ϊ���������,���д��������Ϊ���ݡ����ϡ��߲ˡ�����ֲ��ȱ�������������������������ԭľ����ľ���������������������(Simberloff,2014),��Щֲ��������ߴ�����֡�ũҵ���������š�����ֲ����������뵽����,�漰���롢ʹ�á������ȶ����λ�����,��ͨ�����Ρ�����仯���������õ����ض���ٽ�����ֲ����ɢ(MacDougall et al.,2005),�����̬�ƻ��������������ȷ�����ӱ���ֲ����Ȼ���漰������Ϊ���ŵĹ�������������ֲ�・��ʽ����̬Ӱ��,������������̬�ƻ��Ķ����Ѷȡ�����ֲ���γ������Ժ��֮ǰ,���ڱ�������,��֮�伸ʮ�����ϰ����ʱ��ЧӦ(Caplat et al.,2014),�º������ܡ�
����1.2.3 ��ͷ��������ְ����
��������ְ����ҽ�һ���Ӿ�������ֲ�����������,����,����ά������,����������ֻ����ײ�(Spartina alterniflora)������ݡ��ط������Լ����������߹�ͬ����,�������Ѵ��ͳһ�Ĺ���������������ݺ���������ǿ��ͨ���ײݿ�����Ŀ(ISP),ʮ����Ŭ����ȴ���ֱ�Σ����(Rallusobsoletus obsoletus)�ѽ��ײ���Ϊ��Ϣ��,���ƹ������ò���ֹ(Strong et al.,2016)���ָ�IJ��Ź�����ʽ����̬ϵͳ��������ì��,��ͬ���ŵ�Ŀ���ͻʹ����ֲ��������ӻ���,����,�ڷ���,���ܱ���Ϊ�߶�������ľ,��һЩ�����������ڻ����ƹ���ֲ�̻�(Robinia pseudoacacia)(Dickie et al.,2014)������Ȥ����,����ũ������(ICRAF)�ڷ���ͬʱ�����ͷ�����ֲľ������(Prosopis spp.)ֲ��(Low,2012)��
����1.3 ����������Ŀ�겢����ȫһ��
����ֲ�����ֲ������ڸ���Ӱ��,��������Ȼ�������ı�ľ�����,����ֲ�����̬ϵͳ���������ͬ�����л���Ӱ��,������ֲ��ܿ����������������̬ЧӦ(�羭�ü�ֵ�����ۼ�ֵ��),ֻ����ͼ�ھ����ܴ�������������������ֲ��,�������ֹ�������ᡢ���������ȹ���Ŀ���ͻ(��3)��
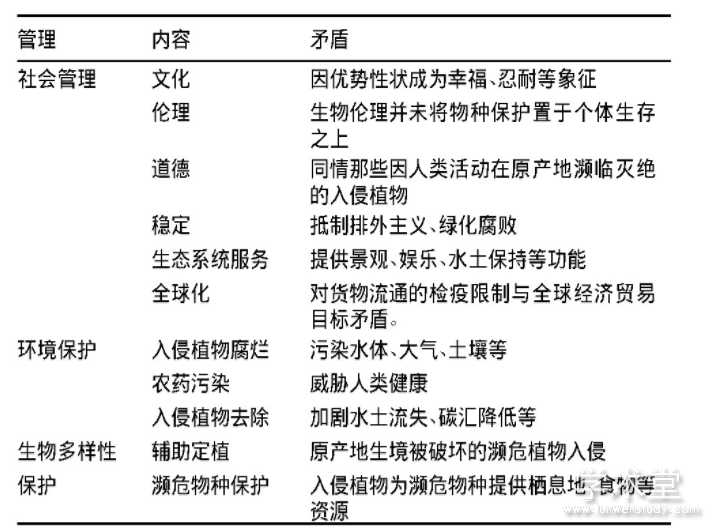
����1.3.1 ��������Ŀ��ì��
���������е�����ֲ����Ϊ��������,��Ϊ�ܱ����������ַ�ֳ����Դ,ͬʱҲ��Ϊ������,Ϊ����������ۡ����ֵ���̬������(McLean et al.,2017),���ڶ�“�̻�����”�����Ǽ�������̬����,��������������ȥ�������е�����ֲ��(��С����,2017)���ܶ��������������ֲ����Ϊ������־,����,�ϷDZ�����������һ���������(the Jacaranda city),����͢�ɺ�������������(Pinus spp.)����(Dickie et al.,2014)������,����ʱ��ϳ�������ֲ��,�������������������������Ե�������״,���뵱���Ļ���,����,����˹��(Cosmos bipinnata)��Ϊ“��ɣ��”(�Ҹ���)(������,2017)�����ڲ�����ͬ������,������ԭ���ر�������������ֵش������ڵ�����ֲ��,������Ұĵ��(Miconia calvescens)��������(Pinus radiata)��,���������������Է���“������ֲ”,�����ڸ����Ϻ���֧�ָ�������ֲ��(Caplat et al.,2014)����Щ���ص�������ֲ��������ܶ���(������̬ѧ��)��Ϊ��“�����ɴ�����(environmental Nazism)”��“��������(xenophobia)”�����Ե��ơ�
������3 ����ֲ�����Ŀ������������Ŀ���ì��
����1.3.2 ����̬��������Ŀ��ì��
����������Ȼ�������ı�ľ�����,�������������ܴ�����ֲ���л���(Tassin et al.,2015),�����ȴ��Ĵ�����,����ֲ������(Cinnamomum camphora)�ӳ��˷۶����(Ptilinopus regina)ʳ����Դ�Ĺ�Ӧ����(Neilan et al.,2006),�����������,������ҪȨ������ֲ���������������Ա���(Vitule et al.,2012)������,���ֹ��������л����������ˮ�����Ⱦ,��ֲ�����ֵ���̬����ת��Ϊ�������⡣�ŻԵ�(2011)���鷢��,��20%��70%�ij��ݼ��������������,�Ӷ�ʹ����������ˮ�ܵ���Ⱦ;de Winton��(2013)�о�����,���������д�Լ��һ��ijɱ�����������ʹ����ջ������ֲ��,Ϊ���ͷ���,�ո������ֲ������ѷ�,�丯�ÿ��ܲ���������������ˮ����Ⱦ�����������������ز��ŵ�����,���ڶԻ�����ȫ�Ŀ���,�ܶ��������ʹ�òݸ�좵�ũҩ��������ˮ��ֲ��(Hussner et al.,2017),��������ֲ��Ѹ����ɢʱ,��ѧ������Ψһ���е�ѡ����,�ܶ���������������������������Ͽ�����̬λ���Ͽ�������ٶ�,��ֹ������ʧ������̼���(Proche��et al.,2012),����������ֲ�������ֱ�ӽ�����Щ��̬���ܡ�
����2 ����ֲ����ƹ����еľ���
��������ֲ�����������ѧ��������ƴ�ʩ�����ھ���,ǰ�ڵ��顢�����������ơ�Ͷ����õȶ���Ӱ����Ƴ�Ч������ֲ��Ĺܿ�Ŀ���ǻָ�����������̬ϵͳ����,���ڿ��Ƹ����á�����ֲ������ЧӦ���̷����ֵ�ԭ��,��ʹ�ɹ�����������������ֲ��,�ܿ�Ŀ���Ժ���ʵ�֡�
����2.1 �߰��ɱ�����������Ʋ�����
��������ֲ������ܳɱ��ɷ�Ϊ���ֳɱ�������ɱ������Ƴɱ�,3�ֳɱ��Ӱ��,�������̶ֳȡ�����Ŀ��Ϳ���ʱ���Ӱ��(Hester et al.,2013)������ֲ����Ƴɱ��߰�,�����ո����������ˮ��ֲ��ijɱ���Լ��ÿ����2000��4000��Ԫ,Ϊ�˴ﵽ�ϸߵ�ȥ��Ч��,����ÿ�����3��,���Ʒ��ý��ﵽÿ����6000��12000��Ԫ(de Winton et al.,2013)������,��������ֲ��ɱ�Զ����������ɢ����ɱ�,van Wilgen��(2016)ͳ�Ʊ���,�Ϸ��ڹ�ȥ20�����������ֲ����ܳɱ�ԼΪ5.64���ϷDZ�,ֻ�д���������ʽ�(1.70��26.08���ϷDZ�),���ܸ���ʣ���(4%)����ֲ�����Ƿ���������Ȼ����ά�ָ߰��Ŀ��Ƴɱ�,�����Ǿ��÷���ĵ���,���Ŀ��Ƴɱ�ҲҪ����������Ȼ�����������ҹ��ȸ�ֵ��̬ϵͳ,��������IJ����������ֲ�������ٴ���ɢ����ͷ��,����Ӱ������ֲ����Ƶ�����Ч��(Kerr et al.,2016)��
����2.2 �����������ƿ��Ƴ�Ч
���������������ܵ�������,���ͻ�е���ѽ���ɽ�ػ��ֵ�,������ˮ������,�ӵ���ˮλ���ӵ��ε�Ҳ��Ӱ���еʹ�á���ѧ���ݼ�Ҳ���ܵ��������ӵ�Ӱ��,���ꡢ��ϫ�ȶ��ή����Ũ��,�ӷ���ũҩ��ֲ������������ʱ��Ҳ���ܵ�����ǿ�ȡ����ٵ�Ӱ��(Kerr et al.,2016)��������Ƽ���Ϊ������,ֻ��������ɸѡ�д����������չ��һ������Ⱥ��ģ,���ܿ�������ֲ��,�һ�����ܵ������Լ���еȻ������ӵ�Ӱ�졣Mahoney��(2017)�о�����,¶��̦ݺ(Oreothlypis luciae)�ͻ�̦ݺ(Setophaga petechia)�Ჶʳ������������������,ʹ��������Ⱥ�����ܵ����ơ���ʹ������Ƽ��ܹ���Ӧ��Щ����,���ڻ����仯,ijЩ���������Ч���Բ�����,����,������Ҷ��(Chrysolina quadrigemina)�������ֹ�Ҷ����(Hypericum perfortum),ɭ�ִ��Ἣ�������˫��Ҷ�״�����ɢ,���»��ַ������������Ч�ʽ���(Swope et al.,2012)��
����2.3 ���ƴ�ʩ������������̬ЧӦ
������Ϊ�����Ǵٽ�����ֲ����ɢ������Ҫ��������,�����Ϳ��ƻ�е��������������,������ҵ��ı�ԭ����ò,����ˮ����ǡ�����ѹʵ������(Dickens et al.,2016)������ֲ��,������ˮ��ֲ��,�ܶ�ܽ��п�¡��ֳ,���䷱ֳ����к�ǿ����������,�и�ջ�����ܿ��ܴٽ�����ֲ�����ɢ�ʹ���(Souza-Alonso et al.,2017)��������Ƽ�����ǰ,ͨ�����������Ⱥ������(centrifugal phylogeny method)�������������Լ��,�Խ��ͷ�Ŀ��Ӱ��,����Ȼ���DZ��������������ȫ���,�����������Ƽ��п��ܳ�Ϊ�µ���������(Hussner et al.,2017)��
�������ڿ��ƾ�������,���ֹ���������Ա����������ֱ��Ӱ��,����,Ortega��(2011)ʹ����Ҷ���ݼ�����������ʸ������ֲ��Centaurea stoebeʱ,������Ҷֲ��ͬ���ܵ����ơ�������ѧ���ݼ����ᶾ����������(���µ�,2016),����Эͬ��������,����������Ⱥ������Ȼ�Ա���ֲ��Ӱ�������,��ʳ�����ܽ�������ֲ��������ͷŻ�õ�����,���֧���˱���ֲ��(Smith et al.,2015),Ȼ�����ֿ��Ʋ�������̬����,�ή�ͱ�����ʳ����ķ�ȡ�����,Carter��(2017)�о�����,ͭ����(Agkistrodon contortrix)��������Ϣ������ֲ���������,�����ڹ�������,��Щ�ط���ͭ�����������ո��ߡ�
����2.4 ���Ʋ���������������ЧӦ
��������ֲ���������ֻ�Ǽ��ھ����ܴ�������������������ֲ��,����������ЧӦ,��̬ϵͳ������������ֲ��Ŀ��ƶ��ָ�(Corbin et al.,2012)������,����ֲ���뱾��ֲ����������ӽ�Ӱ�챾��ֲ���Ŵ�������,Bleeker��(2007)����,�¹���17�ֱ���ֲ����������ֲ���ӽ�������Զ��˥��(outbreeding depression),8��ֲ�������ܻ�����(gene introgression)Ӱ�졣���,����ֲ���е�“��̬ϵͳ����ʦ”(ecosystem engineer)�ı���������ˮ�ĵ���״̬,������ܶȵ�����«έ(Phragmites australis)��ͨ������ˮ���ٶȺ���ɳ�������ı�ӵ�(Kessler et al.,2011),���ֱ��ı��������Ȼ����������ֲ��ȥ�����ָ�������,����ֲ����ͨ����ϵ������ı�������������Ⱥ��,�ƻ�ԭ��������-ֲ�ﹲ����ϵ,���ܸı�����pHֵ���л��ʡ�Ӫ���ɷֵ�(ȫ������,2016),��ʹȥ������ֲ��,�ı�������������ʺ����ﻷ��������������ֲ��(Magnoli et al.,2013)�����,һ��ֲ��ӵ���ֲ������ʧ,�������ӿ�(seed bank)�е�����������ʹ����һ��ʱ���ڴ���,����ֲ���ܹ��ı����ӿ����,�������ӿ��Ⱥ��ָ���“����ЧӦ”(Gioria et al.,2012)��
����2.5 �������������̷�����
��������ֲ����ƺ�����������̷�����,Pearson��(2016)������ֲ����ƽ�����ͳ��,���ֳ���29%���ƹ����ٽ��̷����֡���ǰ,�����������ѳ�ֲ�����ֵij�̬,��Ϊ�̷����ֵ춨����,Ⱥ��������ֲ��֮���Լ�����ֲ���뱾��ֲ��֮��ͨ�����ֳ��ּ侺����ϵ(Kuebbing et al.,2015),�����������ֱ�Ȼ���ͷű���ֲ�P��������ֲ��,������ֲ������ͨ����ԴЧӦ������ͷŵ�ԭ��,���뱾��ֲ�ᄎ���л������(����ҫ��,2018a),����,�ɴ����ݲ�������������������Ʒ������������齬���(Euphorbia esula),Butler��(2010)���ڼ�����,�������������������Ⱥ��������ֲ�����ֲ������ЧӦҲ��ٽ��̷�����,����ֲ�����ܵ��Ŵ������Խ��͡�������ϵ�ƻ�����ʧ,�����ڲ���������Ŀ������ֲ��ȥ�����ָ�,��ȱ���ֲ��,��������ֲ���ܵ���Ӱ����ȻҪС�öࡣ
����3 ȫ��仯��ֲ�����ֹܿص�Ӱ��
��������ֲ�ﲻ��������Ⱥ��仯��“˾��”Ҳ�Ǵ��ȫ��ЧӦ��������ɢ��“�ÿ�”(Waller et al.,2016),���������ظ��DZ仯��������,����ֲ��ͨ��������̬��ʧ������ϵͳ����,ά����̬�����ܡ�
����3.1 ȫ��仯�ٽ�ֲ������,���ӹܿ��Ѷ�
�����������ֻ�Ӿ��������������ɥʧ����̬ϵͳ�������������ʻ���ȫ��仯�������ӵĽ���(���Է���,2010),��һ������������ͨ����Ӫ������ȫ������仯��Ѹ�ٸı���Ȼ����,��Щ�仯ʹ����ֲ�ﲻ��������Ⱥ��仯��“˾��”Ҳ�Ǵ��ȫ��ЧӦ��������ɢ��“�ÿ�”(Waller et al.,2016)����Ŀ�ĵ���״ɸѡ�Ͷ����������������ֲ������ֿ���Դ,����������״�Ŀ����Ը���(Buckley et al.,2016),��ȫ��仯����Ӧ�ϱ���ֲ�����,Lee��(2016)�о�����,��ƽ�����ߡ����������ζ����������ڻ����ײ�����;Mozdzer��(2016)�о�����,CO2Ũ����������������«έ�ĸ���,��ͨ����̬λ����ٽ���ֳ����ɢ;Zhang��(2016)����,CO2Ũ�����ߡ��������ܹ��������ޱ�ʾ�(Mikania micrantha)�ͷɻ���(Chromolaena odorata)������ʺ�ϵ�����Ч�ʡ�
����3.2 ȫ��仯�ı��������,���ܿػ�����
���������������漰��̬����������ᡢ���õȶ������,ȫ��仯����ʹ���������ֲ��Ĺ�����ı�,������������ֲ��ܿصı�Ҫ�Ժͻ����ԡ�����ֲ��һ���浼����̬ϵͳ������ɥʧ(Vaz,2017),��һ����,����ֲ����Ϊ��̬ϵͳ����ɲ���,�������̬ϵͳ����������ظ��DZ仯�Ŀ����Ժ���Ӧ��(Pyke et al.,2008)����Ϊ��̬ϵͳ�Ļ���,����ֲ��Ķ����ԡ���ȡ��ʺ϶ȵ���ȫ����ٱ仯��ֱ��Ӱ��,������ʳ�������¶��ϵı��������������ظ��DZ仯��������,����ֲ�����ͨ��������̬��ʧ������ϵͳ����,ά��̼���桢Ӫ��ѭ������̬����(Mascaro et al.,2012),����,����Լ��,���֖x��(Mangifera indica)��������(Syzygium)����,������άϵ���غֺ���(Eulemur fulvus)����������������(Tonnabel et al.,2011)��
����4 չ��
������Ӧ�ò�������,�������Ƕ��������ֵ��о��������,���յ��о��ɹ���Ҫ����ʵ��Ӧ��,����Ϊ�����ж����������ߵ����ݡ����ڵ�ǰ����ֲ��������ڵ�����,������ǿ���·�����о�:
����(1)��ǿ����ֲ����̬ЧӦ�о�,̽Ѱ����ֲ��Ա�־������(��ϧ����Σ��������ֲ���)��Ӱ��,����������Ӻ���֧�����ȡ���ȫ��仯������,����ͬ�߶���������ֲ����̬ЧӦ�о�,�ḻ���Ƕ�����ֲ��–��������–ȫ��仯��ϵ������,Ϊ��ǿ����ֲ������ṩ���ڻ�����
����(2)��ǿ��������ֲ���ָ��о�����ʹȥ��Ŀ������ֲ��,������������ЧӦ,����Ⱥ���ؽ������ܵ�����,���,Ӧ�ڿ������������ӱ���ֲ��,������ָ�������Ϊ���ֿ��Ƶ�һ����,��ɸѡ��Ѹ��ռ�ݿ�����̬λ�ı������ֻ������,��ȷ�����Ʒ�ʽ�Ա������ֵĸ���Ӱ�졣
����(3)���д�ʱ�ճ߶ȹܿ��о�������ֲ�����������,�о�ʱ�������������������Ȳ���,��߹�����Ч��;�о����ش�С��ͬ,�ó��Ľ��ۿ��ܴ��ڲ���������ȫ�෴��ʵ����,���ǵ����Ρ����õ�����,С�߶ȿ�������ijɹ���������Ӧ���ھ��۳߶�(���˹����ݡ����յ�),���,Ӧ��ȫ��仯������,��ǿ��ʱ�ճ߶�����ֲ������о�,Ϊ����ֲ������ṩ��ѧ���ݡ�
����(4)��ǿ����ֲ��ܿؾ����о���ֲ�����ֲ������ڸ���Ӱ��,������ʶ������ֲ�ﱾ��Ҳ����̬ϵͳ��ɳɷ�,����ֲ���������ͳ����������ֶԱ�����Ȼ/�����̬ϵͳ�Ļ���������Ӱ��,���������Ļ�����ֵ����ܿسɱ����������,��������ǰ���ֲ�Ȩ�����ֹ�������̬�������������ij�ͻ,ͳ��ܿسɱ�����̬��ʧ,���ڿ����볤�ڿ���,�������뷶Χ����,�ص������ȫ����Ƶ�,ͨ���ɱ�����ɸѡ��ѿ��Ʒ���,�Ծ����ܵ��������ֲ��ܿ�Ч�ʡ�
���������
����[]���Է�,ŷ����,л����,��.2010.ȫ��仯��Ҫ���̶Ժ�����̬ϵͳ�������ֵ�Ӱ��.���������,6(6):605-614.
����[]����ͤ,���,ʯ����,��.2012.��ʮ���й����������о���չ.���������,5(20):581-611.
����[]����,���,�Ʊ�,��.2016.��ѧ���ݼ���ũ����̬ϵͳҰ��ֲ������Ե�Ӱ��.���������,24(2):228-236.
����[]ȫ����,ë����,�¼Ҷ�,��.2016.��צ�����������������ֶ�������ѧ������ѧ���ʵ�Ӱ��.ֲ��Ӫ�������ѧ��,22(2):437-449.
����[]����ҫ,���Զ,��ΰ��,��.2018a.“Secondary Invasion”���������ֲ��̷�����.��̬ѧ��־,37(12):3780-3786.
����[]����ҫ,���Զ,Īѵǿ,��.2018b.����ֲ����̬ЧӦ����Ӱ�������о���չ.��ȫ�뻷��ѧ��,1(18):375-380.
����[]��С��,������.2014.dz�����̻����̽����еĸ������⼰�Բ�.�ִ���,(7):144-145.
����[]�Ż�,�ƹ�ǿ.2011.���ݼ�����Σ����ԭ������ش�ʩ.�ִ�ũҵ�Ƽ�,(17):174-175.
����[]������.2017.��ɣ����ʲô.�������ѧ,(3):38-41.
����[]Bleeker WA,Schmitz UB,Ristow M.2007.Interspecific hybridisation between alien and native plant species in Germany and its consequences for native biodiversity.Biological Conservation,37:248-254.
����[]Buckley YM,Catford J.2016.Does the biogeographic origin of species matter?Ecological effects of native and non-native species and the use of origin to guide management.Journal of Ecology,104:4-17.
����[]Butler JL,Wacker SD.2010.Lack of native vegetation recovery following biological control of leafy spurge.Rangeland Ecology and Management,63:553-563.
����[]Caplat P,Hui C,Maxwell BD,et al.2014.Cross-scale management strategies for optimal control of trees invading from source plantations.Biological Invasions,16:677-690.
����[]Carter ET,Ravesi MJ,Eads BC,et al.2017.Invasive plant management creates ecological traps for snakes.Biological Invasions,19:443-453.
����[]Catford JA,Baumgartner JB,Vesk PA,et al.2016.Disentangling the four demographic dimensions of species invasiveness.Journal of Ecology,104:1745-1758.
����[]Corbin JD,D’Antonio CM.2012.Gone but not forgotten?Invasive plants’legacies on community and ecosystem properties.Invasive Plant Science and Management,5:117-124.
����[]Courchamp F,Fournier A,Bellard C,et al.2017.Invasion Biology:Specific problems and possible solutions.Trends in Ecology&Evolution,1:13-22.
����[]deWinton M,Edwards T,Wells R,et al.2013.Review of best management practices for aquatic vegetation control in stormwater ponds,wetlands,and lakes.[EB/OL].[2019-2-18].https://researchcommons.waikato.ac.nz/handle/10289/7999.
����[]Dickens SJM,Mangla S,Preston KL,et al.2016.Embracing variability:Environmental dependence and plant community context in ecological restoration.Restoration Ecology,24:119-127.
����[]Dickie IA,Bennett BM,Burrows LE,et al.2014.Conflicting values:Ecosystem services and invasive tree management.Biological Invasions,16:705-719.
����[]Eviner VT,Garbach K,Baty JH,et al.2012.Measuring the effects of invasive plants on ecosystem services:Challenges and prospects.Invasive Plant Science and Management,5:125-136.
����[]Gioria M,Py?ek P,MoravcováL.2012.Soil seed banks in plant invasions:Promoting species invasiveness and longterm impact on plant community dynamics.Preslia,84:327-350.
����[]Hester SM,Cacho OJ,Dane Panetta F,et al.2013.Economic aspects of post-border weed risk management.Diversity and Distributions,19:580-589.
����[]Hirsch H,Gallien L,Impson FAC,et al.2017.Unresolved native range taxonomy complicates inferences in invasion ecology:Acacia dealbata Link as an example.Biological Invasions,19:1715-1722.
����[]Hussner A,Stiers I,Verhofstad MJJM,et al.2017.Management and control methods of invasive alien freshwater aquatic plants:A review.Aquatic Botany,136:112-137.
����[]Hyndman RJ,Mesgaran MB,Cousens RD.2015.Statistical issues with using herbarium data for the estimation of invasion lag-phases.Biololical Invasions,17:3371-3381.
����[]Kerr DW,Hogle IB,Ort BS,et al.2016.A review of 15 years of Spartina management in the San Francisco Estuary.Biological Invasions,18:2247-2266.
����[]Kessler AC,Merchant JW,Allen CR,et al.2011.Impacts of invasive plants on Sandhill Crane(Grus canadensis)roosting habitat.Invasive Plant Science and Management,4:369-377.
����[]Kettenring KM,Adams CR.2011.Lessons learned from invasive plant control experiments:A systematic review and metaanalysis.Journal of Applied Ecology,48:970-979.
����[]Kleunen MV,Dawson W,Essl F,et al.2015.Global exchange and accumulation of non-native plants.Nature,525:100-107.
����[]Kuebbing SE,Nu?ez MA.2015.Negative,neutral,and positive interactions among nonnative plants:Patterns,processes,and management implications.Global Change Biology,21:926-934.
����[]Lee AK,Ayres DR,Pakenham-Walsh MR,et al.2016.Responses to salinity of Spartina hybrids formed in San Francisco Bay,California(S.alterniflora×foliosa and S.densiflora×foliosa).Biological Invasions,18:2207-2219
����[]Low T.2012.Australian acacias:Weeds or useful trees.Biological Invasions,14:2217-2227.
����[]MacDougall AS,Turkington R.2005.Are invasive species the drivers or passengers of change in degraded ecosystems.Ecology,86:42-55.
����[]Magnoli SM,Kleinhesselink AR,Cushman JH.2013.Responses to invasion and invader removal differ between native and exotic plant groups in a coastal dune.Oecologia,173:1521-1530.
����[]Mahoney SM,Theimer TC,Johnson MJ,et al.2017.Similar dietary but different numerical responses to nonnative tamarisk(Tamarix spp.)by two native warblers.Biological Invasions,19:1935-1950.
����[]Mascaro J,Hughes RF,Schnitzer SA.2012.Novel forests maintain ecosystem processes after the decline of native tree species.Ecological Monographs,82:221-238.
����[]Mason NWH,Palmer DJ,Vetrova V,et al.2017.Accentuating the positive while eliminating the negative of alien tree invasions:A multiple ecosystem services approach to prioritising control efforts.Biological Invasions,19:1181-1195.
����[]McLean P,Gallien L,Wilson JRU,et al.2017.Small urban centres as launching sites for plant invasions in natural areas:Insights from South Africa.Biological Invasions,19:3541-3555.
����[]Mozdzer TJ,Langley JA,Mueller P,et al.2016.Deep rooting and global change facilitate spread of invasive grass.Biological Invasions,18:2619-2631.
����[]Nackley LL,West AG,Skowno AL,et al.2017.The nebulous ecology of native invasions.Trends in Ecology&Evolution,32:814-824.
����[]Neilan W,Catterall CP,Kanowski J,et al.2006.Do frugivorous birds assist rainforest succession in weed dominated oldfield regrowth of subtropical Australia.Biological Conservation,3:393-407.
����[]Otega YK,Pearson DE.2011.Long-term effects of weed control with picloram along a gradient of spotted knapweed invasion.Rangeland Ecology and Management,64:67-77.
����[]Pearson DE,Ortega YK,Runyon JB,et al.2016.Secondary invasion:The bane of weed management.Biological Conservation,197:8-17.
����[]Proche,Wilson JRU,Richardson DM,et al.2012.Native and naturalized range size in Pinus:Relative importance of biogeography,introduction effort and species traits.Global Ecology and Biogeography,21:513-523.
����[]Pyke CR,Thomas R,Porter RD,et al.2008.Current practices and future opportunities for policy on climate change and invasive species.Conservation Biology,22:585-592.
����[]Py?ek P,Pergl J,Essl F,et al.2017.Naturalized alien flora of the world:Species diversity,taxonomic and phylogenetic patterns,geographic distribution and global hotspots of plant invasion.Preslia,89:203-274.
����[]Simberloff D.2014.Biological invasions:What’s worth fighting and what can be won.Ecological Engineering,65:112-121.
����[]Smith LM,Schmitz OJ.2015.Invasive plants may promote predatormediated feedback that inhibits further invasion.Ecology and Evolution,5:2411-2419.
����[]Souza-Alonso P,Gonz?lez L.2017.Don’t leave me behind:Viability of vegetative propagules of the clonal invasive Carpobrotus edulis and implications for plant management.Biological Invasions,19:2171-2183.
����[]Strong DR,Ayres DA.2016.Control and consequences of Spartina spp.invasions with focus upon San Francisco Bay.Biological Invasions,18:2237-2246.
����[]Swope SM,Satterthwaite WH.2012.Variable effects of a generalist parasitoid on a biocontrol seed predator and its target weed.Ecological Applications,22:20-34.
����[]Tassin J,Kull CA.2015.Facing the broader dimensions of biological invasions.Land Use Policy,42:165-169.
����[]Tonnabel J,Tattersall I,Simmen B,et al.2011.Decline and demographic changes in the population of the near threatened brown lemur eulemur fulvus on mayotte.Oryx,45:608-614.
����[]Touza J,Pérez-alonso A,Chas-amil ML,et al.2014.Explaining the rank order of invasive plants by stakeholder groups.Ecological Economics,105:330-341.
����[]van Wilgen BW,Fill JM,Baard J,et al.2016.Historical costs and projected future scenarios for the management of invasive alien plants in protected areas in the Cape Floristic Region.Biological Conservation,200:168-177.
����[]Vaz AS,Kueffer C,Kull CA,et al.2017.Integrating ecosystem services and disservices:Insights from plant invasions.Ecosystem Services,23:94-107.
����[]Vitule JRS,Freire CA,Vazquez DP,et al.2012.Revisiting the potential conservation value of non-native species.Conservation Biology,26:1153-1155.
����[]Waller DM,Mudrak EL,Amatangelo KL,et al.2016.Do associations between native and invasive plants provide signals of invasive impacts.Biological Invasions,18:3465-3480.
����[]Zhang L,Chen X,Wen D.2016.Interactive effects of rising CO2and elevated nitrogen and phosphorus on nitrogen allocation in invasive weeds Mikania micrantha and Chromolaena odorata.Biological Invasions,18:1391-1407.