
花鲈(Lateolabrax japonicus)属鲈形目, 科,花鲈属,是近年来兴起的名优水产养殖品种,因其肉质细嫩,营养丰富,深受消费者喜爱。近年来我国花鲈养殖规模不断扩大,在集约化、规模化、养殖环境恶化等条件下常有病害发生,严重制约花鲈养殖业的发展。
在应用化学药物对水生动物疾病进行防控的过程中,常带来耐药性、药物残留及水体污染等问题。近年来,免疫增强剂的开发和应用为水生动物疾病防控提供了一个新的方向[1]。
β-葡聚糖是一种增强免疫活性的多糖,广泛存在于各种植物和真菌中,具有抗氧化、改善血脂和肠道功能的作用[2-7]。
研究表明,饲料中添加β-葡聚糖能显着提高鱼虾免疫力,从而提高其抗病力[8-9]。本实验室前期研究结果表明,在饲料中添加适量β-葡聚糖能够促进花鲈的生长性能,但关于β-葡聚糖对花鲈免疫能力和抗氧化功能的影响尚未见报道。因此,笔者进一步研究了饲料中添加不同剂量β-葡聚糖对花鲈免疫和抗氧化指标的影响,以便为β-葡聚糖在花鲈养殖上的应用提供更丰富的理论依据。
1 材料与方法
1.1 试验饲料
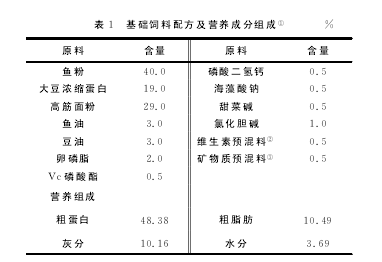
以鱼粉、大豆浓缩蛋白为蛋白源,鱼油、豆油为脂肪源,高筋面粉为糖源,配制半纯化基础饲料(对照饲料),其基础饲料配方及营养成分组成见表1。
在基础 饲 料 中 分 别 添 加200、400、600、800、1000mg/kgβ-葡聚糖(啤酒酵母提取物,纯度>90.0%)配制5种试验饲料。原料经粉碎过40目筛,按配方准确称取逐级混匀后,用SLX-80型双螺杆挤压机制成2.0mm的颗粒饲料,55 ℃下烘干,自然冷却后放入密封袋中,置于冰箱中-20℃保存备用。【1】
1.2 试验鱼及饲养管理
试验用花鲈购自福建诏安县英港育苗场。试验鱼先在水泥池中投喂商品饲料暂养至约8g。养殖试验在广东省农科院动物科学研究所水产实验室室内循环养殖系统的养殖桶(体积约300L)内进行。选取体质量为(8.35±0.17)g的花鲈,随机分成6组,每组4个重复,每个重复30尾鱼。试验开始前用基础饲料驯养1周,待其稳定后,分别饲喂对照饲料和5种试验饲料。日投喂2次(8:30和18:00),日投喂量(以干物质计算)为体质量的4%~6%,试验周期6周。养殖期间,24h供氧,溶氧量>7mg/L,水温为28~30.9 ℃,盐度1,pH 7.5~8.0,氨氮<0.1mg/L。每周换水2次,每次换水量为总水量的1/3。
42d的饲养试验结束后,在原分组的基础上,从每个重复随机取14尾健康花鲈,采用氯化铵进行应激试验,每组4个重复。向水体(体积约300L)中加入氯化铵母液,使非离子氨质量浓度达到3.29mg/L,同时调节水体pH为7.76,应激期间水体pH值用水质监测仪测定。观察花鲈的发病和死亡情况,统计死亡尾数,计算96h内的累计死亡率和相对保护率。非离子氨按下式计算:NH3=[NH4++NH3]/[10(pKa-pH)+1]式中,pKa=0.09018+2729.92/T(T为开氏温度,T=273+t℃)
1.3 样品采集和指标分析
养殖试验结束,禁食24h后取样。每个重复随机取8尾鱼,用1mL注射器自尾静脉取血,血液采集后4℃静置4h,于4℃条件下3500r/min离心10min,取上清液分装,将血清置于冰箱中-80 ℃保存备用。
1.3.1免疫指标测定免疫球蛋白M和补体3含量采用生物素双抗体夹心酶联免疫吸附法测定;碱性磷酸酶活性采用磷酸苯二钠比色法测定;酸性磷酸酶活性采用分光光度法 测 定;一 氧 化 氮 含 量 采 用 硝 酸 还 原 酶 法测定。
1.3.2抗氧化指标测定总抗氧化能力采用比色法测定;超氧化物歧化酶活性采用羟胺法测定;过氧化氢酶活性采用可见分光光度法测定;丙二醛含量采用TBA法测定。
试剂盒均购自南京建成生物工程研究所,具体测定方法参照试剂盒所附说明书进行。
1.4 数据统计
采用SPSS 17.0版软件进行数据统计和分析,试验数据用平均值±标准差表示,显着性水平用P<0.05表示,先对数据作单因子方差分析,若处理间有显着差异,再作Duncan′s多重比较。
2 结果与分析
2.1 饲料中添加β-葡聚糖对花鲈免疫指标的影响
饲料中添加β-葡聚糖对花鲈免疫指标的影响见表2。由表2可见,随着饲料中β-葡聚糖添加量的增加,各免疫指标呈先升后降的趋势,均在400mg/kg组达到最大值。
400mg/kg组和600mg/kg组免疫球 蛋 白M含 量 显 着 高 于 其 他 各 组 (P<0.05)。各添加组补体3的含量显着高于对照组(P<0.05);400mg/kg、600mg/kg和800mg/kg组血清碱性磷酸酶活性显着高于对照组及200mg/kg、1000mg/kg组(P<0.05)。添加量为400mg/kg时,血清酸性磷酸酶活性显着高于对照组和其他各添加组(P<0.05)。
200 mg/kg、400 mg/kg和600mg/kg组一氧化氮含量显着高于对照组及800mg/kg、1000mg/kg组(P<0.05)。
2.2 饲料中添加β-葡聚糖对花鲈抗氧化能力的影响
饲料中添加β-葡聚糖对花鲈抗氧化指标的影响见表3。随着β-葡聚糖添加量的增加,血清超氧化物歧化酶活性呈先升后降趋势,200mg/kg组活性最高,显着高于对照组及800mg/kg、1000mg/kg组(P<0.05)。各组之间血清过氧化氢酶活性差异不显着(P>0.05)。与对照组相比,饲料中添加400mg/kg的β-葡聚糖可以显着提高花鲈总抗氧化能力(P<0.05)。
400mg/kg、600mg/kg和800mg/kg组的丙二醛含量显着低于对照组(P<0.05)。
2.3 氨氮应激时花鲈血清碱性磷酸酶和酸性磷酸酶活性、总抗氧化能力、丙二醛含量的变化
各组花鲈氨氮应激96h时,血清碱性磷酸酶和酸性磷酸酶活性、总抗氧化能力、丙二醛含量见表4。与对照组相比,400mg/kg组的碱性磷酸酶活性显着升高(P<0.05)。
400mg/kg、600mg/kg组的酸性磷酸酶活性显着高于对照组和其他各添加组(P<0.05)。
400mg/kg、600mg/kg组的总抗氧化能力显着高于对照组和其他各添加组(P<0.05)。
400mg/kg、600mg/kg组的丙二醛含量显着低于对照组和其他各添加组(P<0.05)。
3 讨论
3.1 β-葡聚糖对花鲈免疫功能的影响
研究表明,β-葡聚糖能够提高鱼类的特异性和非特异性免疫功能[10]。硬骨鱼类只有免疫球蛋白M,存在于皮肤、肠黏液、胆汁等,利用抗免疫球蛋白M抗体明确证明硬骨鱼类有独立的淋巴细胞种群,对机体的免疫作用非常重要[11]。分泌型免疫球蛋白M由B细胞分泌,出现在血液和其他体液中,作为免疫效应分子存在。
Misra等[12]发现注射10mg/kg的β-葡聚糖能显着提高露斯塔野鲮(Labeorohita)对嗜水气单胞菌(Aeromonas hydrophila)和爱德 华氏菌(Edwardsiella tarda)的 抗 体 效 价。
Figueras等[13]给大菱鲆(Scophthalmus maximus)注射10μgβ-葡聚糖,35d后显着提高其对海鱼弧菌(Vibrio damsela)的 抗 体 效 价。本 试 验 中,400、600、800mg/kg组花鲈血清免疫球蛋白M含量显着高于对照组,猜测β-葡聚糖可能是通过激活淋巴细胞产生大量的抗体对抗细菌和毒素,从而提高花鲈的抗病力。
补体是存在于血清与组织液中的一组经活化后具有酶活性的蛋白质,补体3是补体系统的主要成分,它可以通过经典或旁路途径激活,补体系统的主要作用是促进吞噬作用。有研究报道,β-葡聚糖能够提高水产动物的补体活性。
Misra等[12]用β-葡聚糖对露斯塔野鲮多次注射可以显着提高鱼体补体活性。另外,用含250mg/kg的β-葡聚糖的饲料饲喂露斯塔野鲮幼鱼42d,试验鱼的补体活性达到最高水平[14]。
Bagni等[15]用含1%的β-葡聚糖饲喂尖吻鲈(Dicentrarchus labrax)15d,发现补体活性显着提高。
Engstad等[16]研究发现,对大西洋鲑(Salmo salar)腹腔注射来源于酿酒酵母的β-1,3/1,6-葡聚糖2~4周后,血浆补体活性增加。本试验研究表明,饲料中添加适量的β-葡聚糖显着提高花鲈血清补体3的含量,这与张辽等[17]研究结果一致,说明β-葡聚糖能够通过增强血细胞的吞噬能力来提高补体3的含量,从而提高花鲈的免疫功能。
但长期高剂量投喂会使血清补体3含量呈现降低的趋势,说明β-葡聚糖的添加量及作用时间会影响花鲈的免疫功能。
碱性磷酸酶是一种在碱性条件下能够将对应底物去磷酸化的酶,一般认为在机体防御过程中起一定作用[18],酸性磷酸酶是一种在酸性条件下催化磷酸单酯水解生成无机磷酸的水解酶,两者都是巨噬细胞溶酶体的标志酶,参与动物体内的免疫活动。牟海津等[19]采用虫草多糖和海藻多糖可显着提高栉孔扇贝(Chlamys farreri)血清酸性磷酸酶、碱性磷酸酶活性。本试验结果显示,饲料中添加适量的β-葡聚糖能够显着提高花鲈血清中碱性磷酸酶和酸性磷酸酶活性,这与沈文英等[20-21]在凡纳滨对 虾 (Litopenaeus vannmei)和 中 国 明 对 虾(Fenneropenaeus chinensis)中的研究结果一致。猜测可能是β-葡聚糖能够激活巨噬细胞,通过吸收、破坏、清 除体内的 病原微 生 物,提 高 花 鲈 的 免 疫功能。
一氧化氮是巨噬细胞的效应分子,对T淋巴细胞和巨噬细胞的细胞因子的分泌有调节功能等[22]。
本试验结果显示,除1000mg/kg组外,其他各添加组血清一氧化氮含量显着高于对照组,这与Ohno等[23-24]的研究结果一致。原因可能是β-葡聚糖能够与巨噬细胞表面受体结合,从而刺激巨噬细胞表面iNOS的表达,诱导其产生大量一氧化氮。
免疫球蛋白M以及免疫相关酶活性的升高意味着β-葡聚糖能够提高花鲈机体的特异性和非特异性免疫能力。而高添加量组的花鲈免疫指标显着下降,说明长期高剂量添加β-葡聚糖使花鲈产生了免疫抑制现象。关于β-葡聚糖提高水产养殖动物免疫 力 的 作 用 机 制 尚 不 够 清 楚,有 待 进 一 步研究。
3.2 β-葡聚糖对花鲈抗氧化能力的影响
超氧化物歧化酶是一类重要的清除氧自由基的抗氧化酶,它能催化超氧自由基使其发生歧化反应,生成氧气和过氧化氢;过氧化氢又在过氧化氢酶的作用下,生成无毒的水和氧气[25]。研究表明,采用间隔投喂和浸泡方式,均可显着提高凡纳滨对虾 超 氧 化 物 歧 化 酶 活 性[26-27]。 在 斑 节 对 虾(Penaeus monodon)[28]和中国明对虾[21]中也有相似的报道。丙二醛作为脂质氧化终产物,会引起蛋白质、核酸等生命大分子的交联聚合,且具有细胞毒性,因此丙二醛含量的高低间接反映机体的过氧化程度。总抗氧化能力既可以反映机体清除氧自由基的能力,也易准确测定其含量,所以总抗氧化能力是反映体内活性氧的较好指标[29],反映了机体非酶抗氧化系统和抗氧化酶系统共同完成抗氧化作用。裴素蕊等[30]在凡纳滨对虾饲料中添加虾青素可显着提高其超氧化物歧化酶和过氧化氢酶活性,显着提高其总抗氧化能力。刘冰等[31]对糖尿病小鼠注射壳寡糖,可显着提高机体总抗氧化能力。
金开山等[32]研究发现,花生四烯酸可以降低糖尿病大鼠血清丙二醛含量,提高其总抗氧化能力。王一娟等[29]在中华绒螯蟹(Eriocheir sinensis)饲料中添加抗菌肽可显着降低组织中丙二醛含量,显着提高肝脏组织的超氧化物歧化酶活性以及总抗氧化能力。本试验结果显示,饲料中添加适量的β-葡聚糖可显着提高花鲈血清超氧化物歧化酶活性、总抗氧化能力,降低丙二醛含量,一定程度上提高血清过氧化氢酶活性,与刘群芳等[33]对凡纳滨对虾研究的结果类似,说明β-葡聚糖在一定程度上可以提高花鲈血清抗氧化酶的活性,清除体内自由基,改善脂质代谢,减轻脂质过氧化反应。
3.3 β-葡聚糖对氨氮应激环境下花鲈抗氧化指标的影响
氨氮在水体中主要以离子铵和非离子氨的形态存在。非离子氨是诱发鱼病的主要因子之一,水体非离子氨随着水体温度、pH等条件改变而改变,长期暴露于高含量氨氮环境中会导致鱼体生长性能下降,摄食率降低[34],代谢功能失调,免疫力下降[35]。在受到氨氮胁迫下,机体内增加的活性氧自由基则会通过超氧化物歧化酶、过氧化氢酶和谷胱甘肽过氧化物酶等抗氧化酶的联合作用来对其进行清除,这也是机体克服不良环境和防止中毒的一种适应性反应[36-37]。孙舰军等[38]研究发现,氨氮含量较高时会使中国明对虾超氧化物歧化酶和过氧化物酶活力较对照组有明显下降。张健[39]的研究表明,β-葡聚糖能够有效提高凡纳滨对虾的抗氨氮应激的能力,改善成活率。高举等[40]研究发现,鲫鱼(Carassius auratus)鳃、肝胰脏、肾、脑组织中碱性磷酸酶和酸性磷酸酶的活性随着铅含量的升高和染毒时间的延长均呈下降趋势。本试验中,与对照组相比,各添加组总抗氧化能力、碱性磷酸酶和酸性磷酸酶活性均升高,丙二醛含量均下降,这说明饲料中添加适量的β-葡聚糖可以提高花鲈的免疫和抗氧化能力,增强机体清除自由基的能力,降低对病原的易感性。
4 结论
饲料中添加适量β-葡聚糖显着提高花鲈血清免疫球蛋白M、补体3含量、碱性磷酸酶、酸性磷酸酶活性及一氧化氮含量,提高花鲈的总抗氧化能力、超氧化物歧化酶和过氧化氢酶活性,降低丙二醛含量,从而提高花鲈的免疫和抗氧化水平。





