
湘西黄牛是湖南省优秀的地方家畜品种,主产于湖南省湘西北地区,耐粗饲,耐热,其肉质富含蛋白质和必需氨基酸,脂肪含量适宜,营养全面。
但是,湘西黄牛体型较小,生长发育缓慢,产肉率较低,严重制约了湘西黄牛产业的经济效益。为提高湘西黄牛生长发育速度和产肉率,同时保持其优良肉质,国内研究者已经对湘西黄牛的微卫星多态性和 MC3R、SST、H–FABP、LPL等功能基因的多态性与生长性能的关联进行了探索,结果表明,上述基因的部分多态性位点与湘西黄牛的生长性状相关。但是,这些位点还不足以进行分子育种,更多的标记位点有待于进一步挖掘。
胰岛素样生长因子 I 受体(IGF–IR)是胰岛素样因子家族(IGFs)的重要受体,通过与 IGF–I 结合,共同介导生长激素(GH)的促生长信号通路,在哺乳动物细胞分化与增殖、胚胎发育、神经发育、骨骼肌和骨骼发育等过程中起着重要的调节作用。
迄今为止,未见湘西黄牛 IGF–IR 基因分子遗传特征与生长性状相关的研究报道。本试验采用 PCR 扩增及 PCR–RFLP 技术,分析了 IGF–IR 基因在湘西黄牛群体中的遗传变异及分子遗传特征,并对多态位点与湘西黄牛的生长性状进行了关联分析,现将结果报道如下。
1、 材料与方法
1.1 材 料
1.1.1 供试牛
在湖南省常德地区随机选择 205 头湘西黄牛母牛,经颈静脉采集每头牛的血液,并对牛进行口齿年龄鉴定和生长性状指标(体重、体高、体斜长和胸围)的测定。
1.1.2 试剂
Taq DNA聚合酶、Taq I限制性内切酶购自宝生物工程(大连)有限公司; DNA快速纯化试剂盒、dNTP混合物、DNA Marker II购自天根生化科技(北京)有限公司;100 bp ladder plus Marker购自北京博凌科为生物科技有限公司。
1.1.3 引物
引物(F:CCCA ATGGATTGATCCTCATGT;R:GCTGTGTAGTTCCC TGGGTT),用于扩增IGF–IR基因序列,包括12号外显子(exon12)部分序列、12号内含子(intron12)全序列及13号外显子(exon13)部分序列预期扩增长度为616 bp。
1.2 方 法
1.2.1 基因组 DNA 提取
颈静脉采集2 mL湘西黄牛血液,离心,收集细胞,加血样裂解液、SDS和蛋白酶K共计1 mL以裂解细胞,并采用等体积的酚/仿抽提牛基因组DNA,TE缓冲液溶解,0.8%琼脂糖凝胶电泳检测基因组DNA的纯度,–20 ℃保存,备用。
1.2.2 PCR 扩增
用205头湘西黄牛样本的DNA为模板分别进行PCR扩增。PCR反应体系(20 μL)为:10×Buffer(含Mg2+) 2.0 μL,10 μmol/L上、下游引物各0.5 μL,10μmol/L dNTPs 0.5 μL,2 U/μL Taq DNA聚合酶0.3μL,DNA1 μL,超纯水15.2 μL。PCR反应条件为:95 ℃预变性5 min;35个循环(94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸40 s);终延伸10 min。产物经0.8%琼脂糖凝胶电泳检测后分别用DNA快速纯化试剂盒纯化。
1.2.3 SNP 筛查
随机挑选4头湘西黄牛的PCR产物纯化样品,送生工生物工程(上海)有限公司进行测序,其序列与已公布的安格斯牛IGF–IR基因序列(GenBank登录号为JQ924783)进行多序列比对,确定湘西黄牛SNP数目、突变类型及SNP的位置。将获得的SNP与现有文献资料和SNP数据库进行比较,以确定湘西黄牛IGF–IR基因的新SNP。
1.2.4 RFLP 与基因分型
根据SNP酶切位点的分析结果,确定基因分型位点及RFLP分析用限制性内切酶,对纯化后的PCR产物进行酶切。酶切体系为:PCR纯化产物3 μL,10×限制性内切酶Buffer 1 μL,0.1% BSA 1 μL,限制性内切酶0.5 μL,灭菌双蒸水4.5 μL,65 ℃水浴8~10 h。酶切产物用4%琼脂糖凝胶电泳,凝胶成像系统拍照。根据条带初步判断基因型,并以初步判断的纯合子个体的基因组DNA为模板分别进行PCR(反应条件同上)。PCR产物纯化后送生工生物工程(上海)有限公司测序,验证基因分型的准确性。
1.2.5 数据统计
用POPGENE1.31软件对基因频率和基因型频率、多态信息含量、群体杂合度、有效等位基因数和Hardy–Weinberg平衡χ2检验进行统计。根据影响牛生长性状的因素,在对基因型与表型性状的关联分析时采用了固定模型:Trait(生长性状)=?(均值)+A(年龄)+G(基因型)+e(随机残差),运用SAS(Statistical analysis system) 8.2 软件GLM程序,分析标记基因型对生长性状的影响,并采用文献[21]的方法估计基因效应。
2、 结果与分析
2.1 IGF–IR 基因的扩增结果
PCR产物的电泳结果如图1所示。结合测序结果可知,PCR扩增产物长度为616 bp,其在IGF–IR基因序列(GenBank登录号为JQ924783)中的位置为第95~710 bp。
2.2 IGF–IR 基因 SNP 分析结果
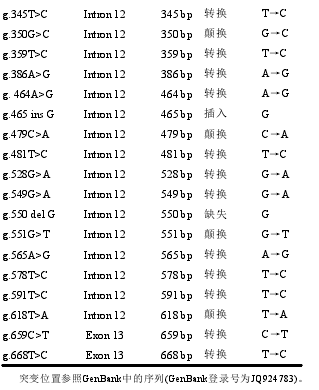
多重比对结果表明,湘西黄牛IGF–IR基因部分序列中存在34个SNPs(表1),其中32个SNPs位于intron 12,2个SNPs位于exon 13;插入突变2个,缺失突变2个,转换突变23个,颠换突变7个;exon 13中的2个SNPs(g.659 C>T和g.668 T>C)均属于酪氨酸的同义突变。34个SNPs中,除了g.551G>T位点有文献报道外,其余33个SNPs均未见相关文献报道,也未有在SNP数据库中收录。

2.3 IGF–IR 基因 g.551G>T 位点的 RFLP 基因分型
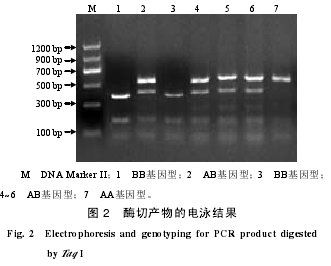
在上述34个SNPs中,仅有g.551G>T位点能被Taq I内切酶所识别,且在其他牛品种上有相关的文献报道。为了能够与其他牛品种群体遗传多态性进行比较,揭示湘西黄牛群体遗传特征,本研究选择g.551G>T位点进行群体遗传多态性分析。PCR产物的TaqI内切酶(识别T?CGA序列)酶切产物的结果(图2)显示,该位点在湘西黄牛群体中共出现3种基因型,分别为AA型(569、47 bp),AB型(569、408、161、47 bp)和BB型(408、161、47 bp)。
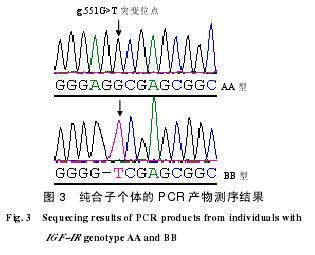
纯合子基因型测序结果(图3)表明,3种基因型的PCR产物的47~50 bp处均存在Taq I限制性内切酶识别位点。
2.4 IGF–IR 基因 g.551G>T 位点的群体遗传特征
IGF–IR 基因 g.551G>T 位点的群体遗传特征统计结果见表 2。3 种基因型分布频率的高低顺序为AB、AA、BB。A 为优势等位基因。多态信息含量为 0.363 7(介于 0.25~0.50)。从杂合度和有效等位基因数可看出,g.551G>T 位点在湘西黄牛群体中处于中度多态性,并且已经达到了 Hardy–Weinberg 平衡状态(P>0.05)。
2.5 IGF–IR 基因 g.551G>T 位点多态性与生长性状的关联分析
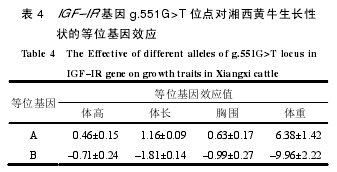
差异显著性检验结果(表 3)表明,湘西黄牛 3种基因型个体的体高、体长、胸围和体重差异均不显著(P>0.05)。等位基因效应结果(表 4)表明,A 等位基因对湘西黄牛的体高、体长、胸围和体重 4 个性状均为正效应,而 B 等位基因均为负效应。综上结果可知,A 等位基因虽然对生长性状均有正效应,但其影响均不显著,因此,从生物统计的角度判断,g.551G>T 位点不适合作为湘西黄牛生长性状的分子标记位点。


3、 讨 论
本试验用文献[12, 17–19]中的引物扩增了湘西黄牛的IGF–IR基因部分序列,长度为616 bp,与GenBank 中收录的安格斯牛的序列长度相同(JQ92478)。但是用同样的引物从郏县红牛、秦川牛、南阳牛、安格斯、荷斯坦牛、西门塔尔杂交牛、安格斯杂交牛和Najdi牛中扩增出的PCR产物长度均为625 bp。
迄今为止,SNP数据库收录了2 233个牛IGF–IR基因的SNPs,可见牛IGF–IR基因具有丰富的遗传多态性。本试验所检测到的34个SNPs均未包含在该数据库中,而仅有g.551G>T有相关文献报道。说明除了g.551G>T之外的33个SNPs均为笔者新发现的SNPs,该结果也说明湘西黄牛群体中IGF–IR基因具有丰富的遗传多态性,可作为揭示湘西黄牛群体遗传特征的候选基因。
本试验PCR–RFLP的结果表明,湘西黄牛与其他牛IGF–IR基因g.551G>T位点的群体遗传多态性有以下差异:酶切片段长度有差异。湘西黄牛在该位点存在3种基因型:AA型(569、47 bp),AB型(569、408、161、47 bp)和BB型(410、170、47 bp),而郏县红牛、秦川牛和南阳牛等品种中的3种基因型则分别为AA型(580、45 bp),AB型(580、410、170、45 bp)和BB型(410、170、45 bp),这种差异是由IGF–IR基因序列长度差异所造成的。
基因型分布频率与部分牛品种有差异。湘西黄牛3种基因型分布频率的高低顺序为AB、AA、BB,与Curi等报道的基因型分布的趋势相同,而Zhang等报道郏县红牛、秦川牛和南阳牛群体中3种基因型分布频率的高低顺序为AA、AB、BB,其中BB型个体数量很少。经分析发现,湘西黄牛AA和AB的基因型频率与上述牛基因型频率分布存在显著差异(P<0.05),而BB基因型频率分布差异不显著(P>0.05)。等位基因频率分布有差异。湘西黄牛A和B等位基因的频率分别为0.609和0.3902。经分析发现,湘西黄牛与Nellore、Canchim牛群体的A和B基因频率分布没有显著差异(P>0.05),而与西门塔尔杂交牛和安格斯杂交牛、郏县红牛、秦川牛和南阳牛、荷斯坦牛和Najdi牛群体的A和B基因频率分布存在显著差异(P<0.05)。群体遗传特征表明,湘西黄牛群体的IGF–IR基因g.551G>T位点处于中度多态,且在群体中达到了Hardy–Weinberg平衡状态(P>0.05),说明该位点不适合用于分子标记辅助育种。
研究表明,1周岁牦牛IGF–IR基因exon 1的SSCP多态位点的EE型个体的胸围、体斜长、体重等显著高于EF和FF型个体(P<0.05),而对于成年牦牛没有影响。Szewczuk等研究表明,牛IGF–IR基因rs41640706/MspI位点(A/G)的GG型个体的断奶重显著高于AG型个体(+5.06 kg)。对于IGF–IR基因g.551G>T位点而言,该位点的RFLP/TaqI不同基因型对南阳牛生长性状影响不显著。上述文献结果显示,IGF–IR基因多态性对不同牛品种(或在同一品种的不同发育阶段)生长性状的影响效果不同。
为了进一步确定IGF–IR基因多态性对湘西黄牛生长性状的关系,笔者进行了g.551G>T位点的关联分析,结果表明,g.551G>T位点的多态性对湘西黄牛群体的体高、体长、胸围和体重均无显著影响(P>0.05),表明g.551G>T位点可能与湘西黄牛的生长性状QTL没有连锁关系,不适合作为湘西黄牛生长性状的分子标记位点。
参考文献:
[1] 于建文.湘西黄牛的特征及选育技术探讨[J].中国畜牧兽医文摘,2013,29(1):76.
[2] 易康乐,李志才,燕海峰,等.湘西黄牛肉质特性研究[J].家畜生态学报,2011,32(5):39–42.
[3] 刘莹莹,肖兵南,田科雄.湘西黄牛的保种与开发利用[J].中国畜禽种业,2008(10):15–17.
[4] 梁斌,李文平,肖兵南,等.微卫星标记测定湘西黄牛杂交牛的遗传多样性及其与生产性能的相关性研究[J].湖南畜牧兽医,2008,30(3):10–13.
[5] 邱志国. 湘西黄牛遗传多样性及微卫星标记与生长发育性状相关性研究[D]. 长沙:湖南农业大学,2007.





