
几十年以前人们已经开始利用微生物来控制害虫。苏云金芽胞杆菌(Bacillus thuringiensis,简称 Bt)是一种独特的细菌,它与许多化合物一起用于防治害虫,并且已经商业化,对农业和环境有重要贡献。尽管其它细菌,包括乳状芽胞杆菌(Bacilluspopilliae)和球形芽胞杆菌(Bacillus sphaericus),也用作微生物杀虫剂,但是它们的杀虫活性谱与 Bt 相比十分有限。重要的是,Bt 对人类是安全的,也是世界上应用最广泛的环境友好型生物杀虫剂。Bt 杀虫基因已经转化到几种主要的作物中,获得了抗虫的转基因植物,进而提供了农业遗传工程模型。
第 9 版的《伯杰氏细菌鉴定手册》将其归属于第二类第十八群,革兰氏阳性,芽胞杆菌属的一个种。
Cry1 类杀虫晶体蛋白对多种鳞翅目害虫具有高效的杀虫活性,是 Bt 杀虫晶体蛋白中研究最为深入的一类,目前杀虫晶体蛋白的作用方式主要是通过 Cry1类蛋白进行研究的。随着cry1类基因在杀虫工程菌和转基因抗虫植物中的广泛应用,抗 Bt 毒素的昆虫已经出现,由于这一威胁,人们希望得到能同时作用在同一害虫上的不同种类的 Bt 蛋白。由于 Cry2Aa蛋白对鳞翅目和双翅目都有活性,所以对 Cry1 类蛋白产生抗性的昆虫,Cry2Aa 蛋白对其也有活性。
Cry2Aa 与 Cry1Aa 的氨基酸序列同源性只有 20%。
然而这两种蛋白的三维空间结构非常相似。这说明 Cry2Aa 蛋白的杀虫机制和许多其它 Cry 蛋白的杀虫机制相似。所以应用 Cry2Aa 来减缓抗性昆虫的出现是有价值的。人们已经提出交替使用 Cry1A 类和 Cry2A 类蛋白来解决抗性问题。长期以来,新型 Bt 基因的分离、筛选以及培育转 Bt 杀虫基因植物,一直是世界上众多国家研究的热点。因此充分发掘我国 Bt 杀虫基因资源已经成为一项极有价值的工作。为此,本研究从土壤中分离 Bt 菌株并筛选和克隆杀虫基因,以期获得具有较高杀虫活性的新型cry1类和cry2类基因。
1、材料与方法
1.1 材料
1.1.1 菌株与质粒 本研究涉及的菌株和质粒见表1。

1.1.2 培养基 液体 LB 培养基 :1.0% 胰蛋白胨、0.5% 酵母提取物、1.0% 氯化钠,pH7.0。固体 LB培养基:在液体 LB 培养基中加1.3%琼脂。固体1/2LB培养基 :1.0% 胰蛋白胨、0.5% 酵母提取物、1.0%氯化钠、1.3% 琼脂,pH7.0。
1.1.3 试剂 限制性内切酶及连接酶、Pfu购自GiBcol、TaKaRa 公司。dNTP、Taq酶等购自上海生工生物工程公司。分析纯化学试剂均为市售。
1.1.4 试虫 小菜蛾(Plutella xylostella)、棉铃虫(Helicoverpaarmigera)和亚洲玉米螟(Ostriniafurna-calis)来自中国农科院植物保护研究所提供的标准化试虫。
1.1.5 PCR 扩增引物 鉴定引物与全长引物均由上海生工合成。引物序列见表 2。
1.2 方法
1.2.1 晶体形态观察 将 Bt 菌株 V4 在 1/2LB 培养基上培养 48 h 后,将胞晶混合液滴于载玻片上,涂抹均匀,烘干固定,石炭酸复红染液染色 3 min,清水冲洗,100× 油镜进行镜检,石炭酸复红染液配制方法参见文献[11]。显微镜下观察晶体释放后,刮取培养物 100 mg 用灭菌蒸馏水洗 3-4 遍,悬浮于1 mL 无菌水中,将芽胞和晶体混合液滴于玻璃片上,待其干燥,经锇酸固定后酒精梯度脱水,临界点干燥,离子溅射喷金(2 nm),HITACHI S -3400N 扫描电镜观察拍照。

1.2.2cry1Ea基因和cry2Aa基因的鉴定及测序 参照宋福平等方法,通过cry1类全长通用引物L5un3/L3un3、cry2类半长通用引物 S5un2/S3un2,以 Bt 菌株 V4 基因组为模板,利用Taq聚合酶进行PCR 扩增,PCR 反应体系为模板 1 μL,引物对各 1μL,2× Taqmix 10 μL,ddH2O 补至 20 μL。PCR 反应 程 序 :94℃ 8 min ;94℃ 1 min 53℃ 1 min,72℃4 min,30 个循环 ;最后 72℃延伸 10 min。获得的cry1类 PCR 产物进行 PCR-RFLP 酶切分析,随后将PCR 产物回收与 pMD19-T 载体进行连接,连接产物转化大肠杆菌 JM109,筛选出阳性转化子,送于上海生工测序。按照已经发表的cry2Aa基因序列,参照文献设计了扩增全长cry2Aa开放阅读框的引物Q2AaF/Q2AaR,并进行cry2Aa全长基因的扩增,引物序列见表 2。
1.2.3cry1Ea和cry2Aa基因序列测定、诱导表达及SDS-PAGE 分析 以 BtV4 菌株的基因组为模板,利用 KOD 高保真 DNA 聚合酶进行 PCR 扩增。PCR 反应体系为 :模板 1 μL,引物对各 1 μL,MgSO43 μL,dNTPs 5 μL,10×Buffer 5 μL,KOD 酶 1 μL,ddH2O补 至 50 μL。PCR 反应程序 :94℃ 2 min ;98℃ 10s,51℃(cry1Ea)/54℃(cry2Aa)30 s,68℃ 4 min(cry1Ea)/2 min(cry2Aa),30 个循环 ;最后 68℃延伸 10 min。之后参考文献[14]。
1.2.4 Cry1Ea 蛋白纯化和 Western blot 印迹 Cry1Ea蛋白纯化用康为世纪的 Ni-Agarose His 标签包涵体蛋白纯化试剂盒进行纯化。用 Bio-Rad Mini Trans-BlotElectrophoretic Transfer Cell 装置进行 Western blot,封闭液为 5% 脱脂牛奶,一抗为 Mouse Anti-His TagMonoclonal Antibody, 二 抗 为 Peroxidase-ConjugatedAffiniPure Goat Anti-Mouse lgG(H+L)。利用 AEC 酶底物试剂盒进行显色反应。
1.2.5 生物活性测定 采用饲料混合法进行杀虫生物活性测定。将 10 mmol/L Tris-Cl(pH8.0)溶解的粗提蛋白制备成蛋白样品溶液,将粗提蛋白溶液与饲料搅拌混匀,分装于经过消毒的培养皿中,挑选活跃的初孵幼虫接于饲料上,进行生物活性测定,每个处理重复 3 次,小菜蛾和玉米螟每个重复为 30 头试虫,棉铃虫每个重复为 24 头试虫。将 10 mmol/LTris-Cl(pH8.0)溶液作为阴性对照,利用同样的方法处理 Cry1Ac 蛋白作为阳性对照。试虫的饲养条件为 28℃、相对湿度为 70%-80% 的光照培养箱中培养,小菜蛾初孵幼虫饲育 48 h、棉铃虫和玉米螟初孵幼虫饲育 168 h 后调查死、活虫数量。
2、结果
2.1 晶体形态观察
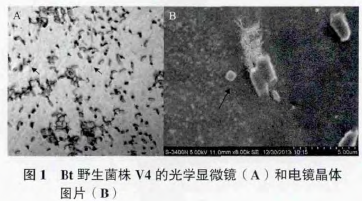
Bt 菌株 V4 产生的晶体形态,如图 1 中光学显微镜下箭头所示,从左到右依次为双锥形晶体和立方形晶体 ;电镜图片中箭头所指,从左到右依次为立方形晶体和双锥形晶体。
2.2cry1Ea基因和cry2Aa基因的鉴定
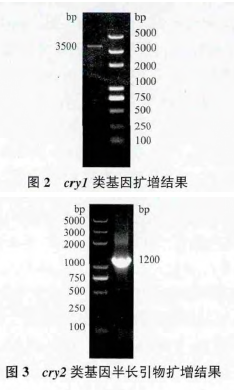
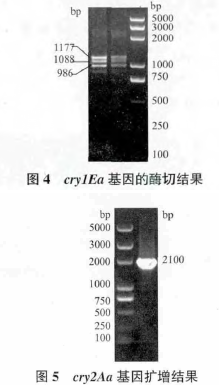
PCR 鉴定发现 Bt V 菌株中含有cry1类cry2类基因,应用引物 L5un3/L3un3 和 S5un2/S3un2 对 BtV4 菌株的基因组进行 PCR 扩增结果,见图 2 和图 3。对cry1类全长基因进行 RFLP 酶切分析,确定该基因为cry1Ea基因(图 4)。参考测序结果,登录 NCBI 进行序列比对,发现该菌株含有cry2Aa基因。利用全长引物 Q2AaF/Q2AaR 扩增 V4 菌株中的cry2Aa基因,PCR 扩增结果见图 5。
2.3cry1Ea和cry2Aa全长基因的克隆和表达
利用全长引物 L5un3/L3un3 和 Q2AaF/Q2AaR扩增 BtV4 菌株中的cry1Ea和cry2Aa基因,将扩增产物分别克隆到 pEB 载体并筛选平末端连接正确的重组质粒 pEB1Ea12 和 pEB2Aa16。对重组质粒pEB1Ea 和 pEB2Aa 基因的测序结果显示,BtV4 菌株中的cry1Ea全长基因大小为 3 507 bp,编码 1 169个氨基酸残基,分子量为 130 kD,并在国际基因库GenBank 中登记,其登录号为 KF601559,由 Btδ-内毒素基因国际命名委员会正式命名为cry1Ea12。该菌株中的cry2Aa全长基因大小为 1 917 bp,编码639 个氨基酸残基,分子量为 60 kD,并在国际基因库 GenBank 中登记,其登录号为 KF667522,由 Btδ-内毒素基因国际命名委员会正式命名为cry2Aa16。
其中,cry1Ea12与cry1Ea11相似性最高,相似度为98.12%,有 65 个碱基差异 ;cry2Aa16与cry2Aa15相似性最高,相似度为 99.47%,有 7 个碱基差异。
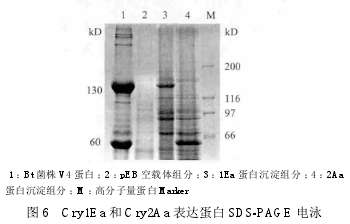
对cry1Ea和cry2Aa基因进行诱导表达,对表达产物进行的 SDS-PAGE 电泳分析结果(图 6)显示,两种表达蛋白被检测均在沉淀组分中,转入重组质粒 pEB1Ea12 的 Rosetta(DE3)菌株中检测到130 kD 的目的蛋白,在转入重组质粒 pEB2Aa16 的Rosetta(DE3)菌株中检测到 60 kD 的目的蛋白,与Bt 菌株 V4 产生的蛋白大小一致。以空质粒 pEB 转入 Rosetta(DE3)菌株作为阴性对照,未发现表达的 130 kD 或 60 kD 的蛋白,以上结果说明在大肠杆菌中成功的表达了上述蛋白。
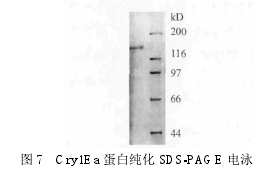
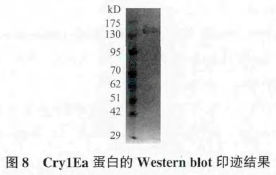
2.4 Cry1Ea蛋白的纯化和Western blot印迹
为了进一步验证cry1Ea基因表达是否正确以及蛋白纯度,对 Cry1Ea 蛋白进行纯化,并进行 SDS-PAGE 蛋白电泳,结果见图 7。Western blot 印迹结果,见图 8。
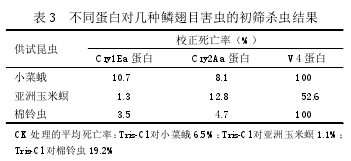
2.5 Cry1Ea和Cry2Aa及BtV4蛋白杀虫活性的测定
提取 Cry1Ea 和 Cry2Aa 表达产物的沉淀组分以及 Bt 菌株 V4 蛋白进行杀虫活性测定,本试验进行了初筛杀虫活性测定。将 3 种蛋白分别设定一个浓度处理,对小菜蛾和棉铃虫为 20 μg/g,对亚洲玉米螟为 50 μg/g,每个处理重复 3 次,并用 10 mmol/LTris-Cl 溶液作为阴性对照。初筛结果(表 3)表明,Cry1Ea 和 Cry2Aa 蛋白对 3 种害虫的杀虫活性很低,但表现出明显的体重抑制,而同时含有该两种蛋白的 Bt 菌株 V4 蛋白的杀虫活性很高。由于供试昆虫数量不足,没有对 Cry1Ea 纯化蛋白进行生物活性测定,也没有做 Cry1Ea 和 Cry2Aa 两种蛋白的混合增效试验,有必要后续继续进行这两个试验。
3、讨论
本研究通过 PCR 扩增反应以及 PCR-RFLP 酶切鉴定,首先确定 Bt 菌株 V4 含有cry1Ea基因,随后发现该菌株也含有cry2Aa基因。Bt 菌株 V4 同时含有cry1Ea和cry2Aa基因,对两种基因表达的蛋白进行杀虫活性测定,结果并不理想。有 3 种可能解释杀虫活性的测定结果 :一是由于碱基的差异导致Cry1Ea12 和 Cry2Aa16 蛋白的空间构象发生改变,致使其杀虫活性明显降低,Cry1Ea 和 Cry2Aa 蛋白对小菜蛾活性很高并且与其它蛋白具有显著的协同增效作用,而 Cry2Aa 与 Cry1A 类蛋白作用于昆虫的受体不同,这能够解释为什么 Cry2A类蛋白能杀死对 Cry1A 类蛋白产生抗性的昆虫,所以 Cry1Ea 蛋白和 Cry2Aa 蛋白有可能具有协同增效作用,极大增加彼此的杀虫活性,但由于供试昆虫数量不足,并没有做两种蛋白的协同增效试验 ;二是有可能存在其它未知基因,表达的蛋白具有很高的杀虫活性 ;三是本试验采用包涵体蛋白进行生物活性测定,但是包涵体蛋白降解速率较快,而生物活性测定时间较长,这可能导致杀虫活性的降低。
此外,Cry2Aa 蛋白对双翅目也有高活性,可以对本试验的两种蛋白进行其它不同害虫的生物活性测定,因此有必要进行后续研究和重复性试验,这对于构建高效工程菌、转抗虫基因植物、解决昆虫对杀虫基因产生抗性等问题是有意义的。
4、结论
以本实验室分离得到的 300 株苏云金芽胞杆菌菌株 DNA 基因组为模板,利用cry1类全长通用引物进行 PCR-RFLP 基因鉴定,发现 Bt 菌株 V4 含有cry1Ea基因,克隆基因全长序列为 3 507 bp,编码 1 169 个氨基酸残基,分子量为 130 kD,并在国际基因库 GenBank 中登记,其登录号为 KF601559,由 Btδ-内毒素基因国际命名委员会正式命名为cry1Ea12。同时发现该菌株还含有cry2Aa基因,克隆基因全长序列为 1 917 bp,编码 639 个氨基酸残基,分子量为 60 kD,并在国际基因库 GenBank 中登记,其登录号为 KF667522,由 Btδ-内毒素基因国际命名委员会正式命名为cry2Aa16。将两种基因在大肠杆菌 Rosetta(DE3)中表达,并表达蛋白进行杀虫活性测定,两种基因表达的蛋白杀虫活性不高,但 Bt菌株 V4 蛋白具有较高的活性。





