
������ũҵ�������ɺ�����ɵ���ʧռũ������ʧ�����ģ��������ϣ���˺��������ũҵ�����е�һ����Ҫ�������没���P���л�����ѳ�Ϊ����ũҩ����Ҫ��ɲ��֡����ƽ�ѿ���˾����£���죭������������������£�����Ŀǰ�������о���Ϊ���롢Ӧ�������㡢��Ӧ����ɹ���ɱ������ԣӣ��������䡡�ף�������������¡�˵ڣ����£�ɱ�澧�嵰���������������Ѿ���¡�ˣ������������� �������� �� �� �� Ŀ �� �࣬ռ�������֡�
�����������ɻ�����һ��ʮ������ģ�����������������뵰��ֻ�У�����ģ���ͬ�ڴ�����ã�������ģ�������ġ��ѹ����ģ��������������ҪΪ�������ɣᡢ�������ɣ⡢�������ɣ�ȣ�������֣����У������ɣ����У����֡��ԣ�����ȴӣ£����ꣴ�������п�¡�˵ڣ����������ɻ���������ɣᣱ����������ʳ�Ŀ���������׳棨�̣������������ᡡ��������������ᣩ���۳�Ŀ��ŷ�����������ϣ�������ᡡ������������ж�ɱ���ԡ��������ֵģ������ɣ������ʳ�Ŀ�����δ���ֻ��ԡ�����Ϊֹ�������ֵģ������ɻ����������ɱ�浰����Ҫ���۳�Ŀҹ��� ���Σ���������壩����Ҷ��ƣ��ԣ�����飭�����壩���˶�ƣ��У�����������壩���ʳ�ĿҶ�ƣ��ã���������������壩�� �� �� �� �� �� ɱ �� �� �ԡ� �� �⣬��������������ڣ£������ж����dz�Ĭ�ģ���������ɣᣱ�����ھ��ꣴ�������о�δ��������
�����������ɻ����Ĭ����Ҫԭ������ǣ������������������������������ڣ��γɣ�������������������������ˣ������ɻ������������ӵ�ȱ����
�������������о����֣����������������˾��н�ǿ��ɱ����Ժ���ԽϹ��ɱ���ף�����������ã�������൰���������ԣ��⽫Ϊ����£�����ɱ����խ�����濹ҩ�Բ��������⣬�ṩ�µı�ѡ������ˣ����������������о�������Ҫ�����������ʵ�ü�ֵ��
�����ǣӣ��DZ�ʵ�������з����Ұ�����꣬���Уãң��ңƣ̣м������У������ɣ�����о���¡�˸û�������������бȶԷ��������ڴ˾��н����˱���ⶨ�������������Լ�����Ҫũҵ����Ļ��ԣ����ָû����С�˶꣨�У��������ᡡ�������������ᣩ���и߶�����
���������Ϻͷ���
��������������
�����������������������������þ������������1.
����

�������������������������̣�Һ�����������ԣ��������塡�����������٣�����������������������������Σ�ã졡������������ȡ����������̣¹�������������Һ��̣��������м��룱��������֬�ۡ�
��������������ø�������Լ��Уã�����Լ������Ϻ����������� �� �� �� ø���ԣ� �ģΣ���������塢�ģΣ����ƣ��磭���������У�������������ˣ�������ԣУ������ṫ˾��
�������������������������ҹ�꣨�ӣ���������ᡡ��������ᣩ���� �� �� ���ȣ����������ᡡ���������ᣩ�� �� �� ҹ �꣨�ԣ�����������ᡡ��飩���ɺӱ�ũҵ��ѧ���没�����������ѧʵ�����ṩ��С�˶꣨�У��������ᡡ�������������ᣩ�ɺӱ�ʡֲ�ﱣ���о����ṩ���Ҳϣ��£��������������飩Ʒ��Ϊݼ��×��£�����ɽ����ͨ���ּ�������˾������Ҷ�ף��У���������ᡡ���������ɼ���Ұ�⡣
��������������
�����������������£���������ȡ���øĽ��ģãԣ��·������μ����ףۣ����ݡ��������ɻ���Уãң��ңƣ̣м������£����������������ת���ӵ�ɸѡ�ο��θ�ƽ�ȷ�����
�����������������˾����ӿ�¡���⡡�ţ�����������ģΣ��� ȡ���ģΣ�ø �С��� �ӡ�ת ������ �� �� �� �ͣӣģӣ��У��ǣż��������ףۣ����ݡ�
�����������������£����ǣӣ������еģ������ɣ����ģУã���������¡�ͱ���ο��ǣ��£����й����ģ������ɣ������������ģ����˺ͣ��������У�������
����

�����ԣǣӣ��������ģΣ�Ϊģ�壬���ø߱���ģΣ��ۺ�ø���УУã�������
�����Уãҷ�Ӧ���������� ��Ԥ���ԣ��������� ����ԣ������������� ���˻𣱣������������죱��������������ѭ������� �����죱���������գУãҲ����Ŀ��Ƭ���ã£���Ȣ�ͣӣ������˫ø�У�����ͬ��ø�д������ģ�ţԣ�����������ӣ�ת���˾��ԣǣ����ԣУãҺ�������ø��ɸѡ������������¡��
�����������������������ɣ�������ͷ���ѡȡ���������¡�������вⶨ���ģΣ����������Ϻ��������﹤�̼�����������˾��ɣ����ãģΣ��ͣ��Ρ��ģΣ����ӣɣӵ������ͣţ��У��ӣ����ݿ�������С�
�����������������������ɣ�����ڴ˾��б��ォ���У������ɣ�ȫ�������������������˾��£̣������ģţ������ԣɣУԣǽ����յ�����ã������ɣ�������������յ������μ��Σ������˾��Ʒ˵����
�����������������ã�����ɣ⵰��ɱ��������Բⶨ����桢����ҹ������ҹ��������ϱ��渲�Ƿ���������ⶨ��������ƽ���ڣ��������С�еײ���Ȼ������μ�̾�Һ����Ϳ�������ϱ��棬��ɣ�ÿ�У���ͷ�Գ棬�ظ����Ρ����ڣ����������������У�ÿ��������һ�λ���������Բ���˳�����룲����Ϊ���������ҲϺ�С�˶���ý�Һ������������갴Ԥ����ƺõ�Ũ�ȣ�����ˮϡ�ͣ�����ˮ��Ϊ���Զ��ա�������ҶƬ��ϴ�����ɣ��ֱ��ڣ£���Һ�н�����ȡ�������ɣ�������ƿ�У�ÿƬҶ������ˮ��ʪ����֬����ҶƬ���ʣ���ÿƿ�ӳ棱��ͷ��ÿ����������ͷ�棬�����ظ��������׳档���룲��������������������ֱ��ڣ��������е��顣����Ҷ�ײμ��Žܵȷ������ⶨ����ã����������ʽ���������ʡ��̣ã����ͣ���������Ӧ�ãУϣ̣���������ó���
����������ͷ���
����������ȫ���������ɣ�������������¡
�����ԣǣӣ����������ģΣ�Ϊģ�壬������������УУã�������ͼ�������پ�������ø�кͲ��������֤���þ��꺬�У������ɣ�Ļ���ͨ���Уã��������ӣǣӣ������¡��ȫ��Ϊ����������⡡�������ɣ�ȫ�������£���Ȣ�ͣӣ���ø�в�����������ţԣ��������У����������������Ϊ�ԣ�ţԣ����ɣ⣨ͼ������
����
�������������вⶨ������
�������ԣ�ţԣ����ɣ���в���õ��������ɣ�����ȫ�� Ƭ �� �� �� �� �� �У��ڣǣ��£���� ¼ �� Ϊ�ţգ����������������ɹ��ʣ£���δ���ڶ��ػ�������ίԱ������Ϊ�������ɣ⣳���Ʋ��˸û����ɣ���������������ɣ��� �� �� �� �� �� �� �� �� �� �� �� ���̣������˿ �� �ᣨ�ӣ���հ��ᣨ�ԣ������ߣ��ֱ�Ϊ�������������������������������������õ���Ԥ�������Ϊ��������������ģᣬ�ȵ��Ϊ��ɡ����������������������Ե��ס�
����ͨ����������Σԣ���������������������õ������ѹ����ģ��֣ã�����ɣ⵰�İ��������д��ڲ��죨����������һ���µģã�����ɣᵰ�ף�������������ʣ£�ɱ�澧�嵰������ίԱ�Ὣ������Ϊ�ã�����ɣ⣳��
����

����ͨ���ãΣã£ɡ��ã��������䡡�ģ������ģ�����������ã�����ɣ⣳���ؽṹ�����������ģ����������ɣζ˵ڣ�����������λ������������������л���ɣ��ģ��������ɵڣ�������������λ������������������л���ɣ��ģ��������ɵڣ�������������λ������������������л���ɣ���ˣ��ã�����ɣ⣳���Ļ���������Ӧλ�ڣζ˵ĵڣ�������������������л�����ͼ������
����

��������ͬԴ��ģ���������ˣã�����ɣ⣳���������ṹ����ʾ��ͼ����
����
�������������������ɣ⣳����ı���
���������Կ�¡���������ԣ�ţԣ����ɣ�ת�����˾��£̣������ģţ����У������յ��������������յ������������Ŀ�ĵ�����Ʒ���Уӣģӣ��У��ǣŷ������ɼ�Լ������ĵ�Ŀ�ĵ������յ�ʱ����ӳ������ױ����� �� �� �� �࣬�� �ڣ�С ʱ �� �� �� ���ɣ�������ͣ�������ģУ�������������������Լռ�ܵ��ģ�������ͬʱ�У�����ģ�ĵ��ײ�������ͼ������
����

����������ɱ��������Բⶨ
�����ɣ�ţԣ��������ģã�����ɣ⣳����С�˶��к�ǿ��ɱ����ԣ��̣ã���ֵΪ��������������μ�磯���̣��������Գ�û�л��ԣ�����������
����
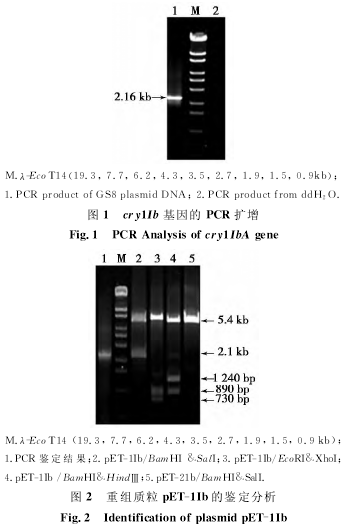
���������������
�������о����ãУãң��ңƣ̣���ϵ����Ұ������ǣӣ��м������������ɣ�����ڴ˻����ϣ��ӣǣӣ��п�¡�ˣ������ɣ�����ڴ˾��гɹ��ı����ˣ�����ĵ����鵰�ס��������ɣ⣳���������ᆳ�ӣģӣ��У��ǣŷ���������Ϊ�����������ﲢ���ǵ�һ�ģ���������������з��֣�ͬһ������£���ͬ����ʼ�����ӹ���һ����ֹ�ӣ������������ڲ�ͬ��ʼ�����Ӵ��������£�Ҳ�п��������ڱ��ﵰ����Ȼ�������£����д��ڽ�һ���о������ã������ɣ⣳����� �� �� �� �� �� �� �� �� �� �� �� �� �� ������ �֣ã�����ɣ⣳������Ҫ���߲˺���С�˶���ʾ�˸߶�������������桢���ҹ�ꡢ����ҹ�ꡢ�ҲϺ�����Ҷ��δ���ֳ�ɱ����ԡ��ݱ��������ڵ��������ṹ�IJ��죬������ĵģã����������ã��������ȣ�������ĵģã�������൰�ײ����ڽ������ԡ���ˣ����о����ֵģ������ɣ⣳�»���Ϊ�ҹ�����ת����������̾������ƣ��ṩ�µĻ�����Դ��Ϊɸѡ���ӻ��Ϳ˷�����������ԵĻ�������ṩ����Ҫ���ϣ�������Ҫ�����ۺ�ʵ�����塣
������������о����֤�����������������Ұ��������ͨ���dz�Ĭ�ģ���������ɣ�����ڹ��ꣴ�������о�û�б������ڣ������ɻ������������һ�����������������ڣ���Ϊ��������������������������ˣ������ɻ������������ӵ�ȱ�������������ڣ£��������ձ���ڣ��ǣ������ɻ����Ĭ����Ҫԭ��
�����ɴ���ɱ�������ɸѡ����Ĵ�ͳ���������ָ������Ĵ��ڡ�ʵ���ϣ��������������㷺�����ڣ£������С������ǵڣ�����¡�Ķ��۳�Ŀ���ʳ�Ŀ�������˫�ػ��ԣ������ɣ�������θ�ƽ�ȿ� ¡ �ģ������ɣ�� ���� �� �� ʵ �� �� �� ¡ �ģ������ɣ⣳�������Ĭ��������������۳�Ŀ�Ⱥ�����кܸߵĻ��ԣ�Ŀǰ��һЩ���������������Ӧ���ڿ���ת����ֲ������ơ��о������������Ϊ��Ҫɱ������ģ£�������ԣ����Ĭ������Դ���ž���о�DZ���������˹�����ѧ���ǵĹ�ע��������Ϊ���£��о��µ��ȵ㡣
�����ο����ף�
�����ۣ��ݣӣ�����桡�ȡ��ţ��ף�����������ȡ��ң��ã�����硡���䡡�ţ����������桡����塡�£���������������������ã�������졡�У�������ǣ��塡���ţ���������ᡡ�����ۣʣݣ��У��㡡�Σ���顡�����䡡�ӣ�飬������������������������������������������
�����ۣ��ݣ�����𣺣������������������飮������������㣮���룯�����壯�Σ���ߣã���룭�����壯�£������������������죮
�����ۣ��ݣã��������塡�Ρ��ģ��ڣ�����ң��ƣ���������ʣ��������죮�ң���飭������桡����塡���������������塡������塡�£��������������������?��������������졡���������졡����������ۣʣݣ��ͣ����������������������������������������������������������������������������
�����ۣ��ݣԣ����ҡ��ʣ��ԣ����������ǣ��ǣ��⣬�������죮�ɣ��������������������������������������桡�ᡡ������졡�£�������������������������ᣭ������������������������졡������������������䡡���������������������ۣʣݣ��ͣ�졡�ͣ�������죬����������������������������������
�����ۣ��ݣã��顡�ӡ��ˣ��ӣ���¡��ӣ��ˣ��硡�š��ͣ��������죮�ã�����硡��桡��������£��������������������������ɣ�������塡�ã�������졡�У�����������ۣʣݣ��ã���ͣ�������죬����������������������������
�����ۣ��ݣӣ���֣��ȣ��䡡�£��ԣ����������ʣ��������죮�£����������졡����������������������������������桡����塡�������������졡����磭����������桡�ã���֣ۣʣݣ���������䡡���䡡�ţ��������������졡�ͣ飭�������������������������������������������������������
�����ۣ��ݣ����������������ӣ��֣���ᡡ�ӡ��ң��ˣ�����С������������죮�ԣ���������������桡�Σ��������ᡡ������������������衡�ᡡ��������塡�������ɣᣵ����塡����������������塡�����������������������ȣ����������������ۣʣݣ��ͣ��������£������磬������������������������������
�����ۣ��ݣӣ��硡�ơ��У��ڣ���硡�ʣ��ǣ��������أ��������죮�ɣ�����������������������ɣ�������塡��������塡��������£�������������������������������䡡���������������������桡�ᡡ������졡�������ɣ������������ۣʣݣ���������䡡���䡡�ţ��������������졡�ͣ�������������������������������������������������������
�����ۣ��ݣţ��������ɡ��ң��ţ�����ᡡ�����У����ͣ��������죮�ͣ�����������䡡�������������졡���������������������桡�ᡡ�ã�����ɣ��������������������������桡����塡���������Σ���������壬�ԣ����������壬�У�����壭������塡���䡡�ã��������������ۣʣݣ���������䡡���䡡�ţ��������������졡�ͣ�������������������������������������������������������
�����ۣ����ݣģ����ȡ��ӣ��ʣ���塡�ӣ�����������ᡡ�£��������죮�ԣ��������������£���������������������� δ����������������������������������������ţ�������ᡡ��������ᡡ�ף����������壬�ᡡ���ᣭ������������������������������ۣʣݣ��ʣ������졡��桡�ɣ������������塡�У�������������������������������������������
�����ۣ����ݣˣ��������ᡡ�ˣ��ף�����ǡ��ף��ͣ������ͣ��������죮�ã�����硡��桡�ᡡ�����֣�������塡�������������졡������������塡������£ᣭ������������������������塡�����֣��������䡡�У�������������������䡡�������������������������������ۣʣݣ��ʡ��£��������죬��������������������������������������
�����ۣ������θ�ƽ���Žܣ��˰��ǣ��ȣ����ƽ�ѿ���˾��������ɻ����Ĭ �� �� �� �ۣʣݣ��� Ȼ �� ѧ �� չ������������������������������������
�����ۣ����ݣԣ�����顡�ӣ��ڣ�����顡�Σ��ʣ����ᣮ�ã�����硡���䡡������������桡�����������������桡�ᡡ������졡�������ɣᣭ������塡����塡������£���������������������������𡡣����������ۣʣݣ��ʣ������졡��桡���������ͣ�����������������������������������������
�����ۣ�������͢�ԣ���Ρ���꽨�㣮���ƽ�ѿ���˾��ǣӣ����������ѧ �� �� �������� �� �� �� �� �ۣʣݣ�ũ ҩ ѧ ѧ ��������������������������������������
�����ۣ����ݣӣ�������롡�ʣ��ƣ���������䡡�š��ƣ��ͣ�������ԣ��ͣ���������������磬���������������������������ۣͣݣ��������������Σ���٣��룺�ã��䡡�ӣ����硡�ȣ����̣�������������У������������
�����ۣ������θ�ƽ���Žܣ��ƴ�P���ȣ����ƽ�ѿ���˾���������Уãң��ңƣ̣м�����ϵ�Ľ����ۣʣݣ��й�ũҵ��ѧ������������������������������������
�����ۣ����ݹ�Ρ������ҹ������ƽ�ѿ���˾��ã��������Ŀ��Լ��������ϵ���о��ۣģݣ����ݣ���ɽ��ѧ������������
�����ۣ����ݣڣ���ʡ��ڣ��ã��ʣ��̣顡�١��أ��������죮�ԣ��������㡡�У������ţ���������硡�ԣ���£���������������������ԣ�����ģ�����ɣ��������ң���������塡�ţ�����������ۣʣݣ��Σ������塡�£�����������������������������������������������������������
�����ۣ������Žܣ��θ�ƽ����ѣ��ȣ����ʳ�Ŀ����߶����£��������������ķ����¡�������о��ۣʣݣ��й�ũҵ��ѧ����������������������������������������
�����ۣ����ݣ�����������ס��ӣ������ͣ�����䡡��桡�ã����������硡����塡�ţ���������壭�����桡���ɣ����������ۣʣݣ��ʡ��ţ���ţ�������죬��������������������������������
�����ۣ������θ�ƽ�����ƽ�ѿ�߸˾���������壱�ͣ�����������о��ۣģݣ��������й�ũҵ��ѧԺ������������
�����ۣ���������Ѧ�����θ�ƽ���ȣ����ƽ�ѿ���˾��£��ã�����ɱ�澧�嵰�����������¡��ɱ����Է����ۣʣݣ�ֲ�ﱣ������������������������������������





