
间套作在没有扩大土地面积的前提下提高了粮食产量,为解决世界人口的温饱问题做出了重要贡献。研究表明,间套作种植能够有效提高资源利用率和粮食产量,增强农业系统的抗风险能力,增加水土保持能力,提高土壤肥力,同时能够抑制病虫草害的发生,是一种基于生物多样性的生态农业范式。在当前全球环境变化加剧大背景下,间套作种植模式的研究越来越受到关注。在我国西南丘陵地区,玉米-大豆带状套作种植近年发展迅速,四川省自 2003 年开始推广以来,种植面积已达 115.1万公顷,农民增加收益 60.65 亿元,显著提高了区域粮食生产能力。
株高不同的作物搭配构成的间套复合群体中,高位作物对低位作物造成的遮荫将会影响到低位作物的生长发育,进而影响到间套复合系统的生产力。国内外在高位作物对低位作物形态发育和生理特征影响方面已有相关报道,不同荫蔽程度、不同作物间均存在较大差异。在玉米-大豆套作模式中,大豆播种于玉米行间,幼苗出土后便生长在玉米荫蔽环境中直至玉米收获,共生期约 60 天左右,玉米荫蔽环境将会导致套作大豆光合有效辐射截获减少,叶片结构发生变化,茎秆变细变长,这将可能伴随着一系列生理生态特性的适应,却尚未见相关报道。本文以耐阴性不同的大豆品种为研究对象,从光合生理生态的适应性角度探讨大豆幼苗对玉米遮荫环境的响应,旨在为大豆耐荫品种选育提供理论依据,同时为玉米-大豆套作技术完善和应用推广提供技术支持。
1、 材料与方法
1.1 研究地点概况
试验于 2012 年在四川农业大学教学科研农场进行,研究区位于四川省雅安市雨城区(29°59′N,103°00′E)。该区域属于亚热带湿润季风气候,年平均气温 16.2 ℃,月最低气温 6.1 ℃,月最高气温 25.4 ℃,无霜期 280—310 d;降水资源丰富,年降雨量 1250—1750 mm;光资源相对不足,属于全国太阳辐射较低的区域,年均日照时数仅 1005 h,太阳总辐射 3750 MJ/m2/y左右。试验地土壤类型为紫色粘土类型,pH值约 7.5,有机质含量 29.8 g/kg,全氮 1.3 g/kg-1,全磷 0.5 g/kg,全钾 26.2 g/kg,速效氮 176.9 mg/kg,速效磷 38.1 mg/kg,速效钾 129.6 mg/kg。
1.2 试验设计
试验采用双因素随机区组设计,每小区面积 6 m×6 m=36 m2,3 次重复,共 12 个处理小区。主因素为种植方式:大豆单作(M)和大豆-玉米套作(RI);副因素为品种:南豆 12(ND12)和南冬抗 022-2(NDK)。在玉米-大豆套作种植模式中,带宽 2 m,玉米品种选用川单 418(株型半紧凑、株高 269 cm),由四川农业大学玉米研究所提供;大豆选择 2 个耐荫性存在差异的品种:南豆 12 和南冬抗 022-2,由四川省南充市农科院提供。玉米于 4 月初播种,采用南-北行向宽窄行种植方式,窄行行距为 40 cm,穴距 19 cm,每穴 1 苗,全田玉米密度为 5.25×104株/hm2;大豆于 6 月初在玉米吐丝期播种 2 行于玉米宽行内,行距为 40 cm,穴距 19 cm,每穴 2 苗,全田密度为 1.05×105株/hm2,大豆、玉米行间距离为 60 cm。大豆单作种植模式中,大豆播种期与套作模式相同,行距 50 cm,穴距 19 cm,每穴 1 苗,密度 1.05×105株/hm2,保证单株的生长空间与套作模式相同。试验以每公顷纯氮 36 kg、P2O563 kg、K2O 52.5 kg 做为大豆底肥施入,其他农艺措施管理同常规大田生产。
1.3 测定指标与方法
1.3.1 大豆生长微气候环境指标测定 选择晴朗天气(分别于大豆出苗后第 30 天、36 天和 43 天)从 6:00—18:00,每间隔 2 h,在大豆幼苗冠层上方(冠层上方10 cm处)利用光量子传感器 LI-190 (LI-COR, Lincoln, USA) 进行入射光合有效辐射(PAR)日动态变化的测定(每小区选择 6 个点),同时利用 HOBO H8 温、湿度数据采集器(Onset Computer Corp,Bourne, USA)进行温度(TEMP)和相对湿度(RH)日变化的测定。
1.3.2 光合速率和叶绿素荧光参数测定 大豆出苗后第 30、36 和 43 天,从 9:00—11:00,每处理小区选择 6 株大豆幼苗的功能叶(倒二叶),利用 LI-6400 便携式气体交换测定系统 (Li-COR, Lincoln, USA)进行瞬时气体交换参数的测定。测定时光照采用人工光源,光强控制在 1000 μmol m-2s-1,叶室内温度为 24—28 ℃,相对湿度 65%—75%。在测定气体交换参数的同时,利用 Mini-PAM 便携式脉冲调制式叶绿素荧光仪(Mini-PAM, Heinz Walz, Effeltrich,Germany) 测定最大荧光 Fm和初始荧光 Fo,测定时将叶片充分的暗适应 30min,每一处理小区选择有代表性的 6 株,测定其功能叶片(倒二叶)。可变荧光 Fv 和 PSⅡ最大光化学量子产量 Fv/Fm计算获得:Fv= Fm–Fo;Fv/Fm= (Fm–F0)/ Fm。
1.3.3 叶片采集和叶绿素含量测定 大豆出苗后第 30、36 和 43 天,每处理小区选择 12 株,取其功能叶(倒二叶)迅速放入液氮中,带回实验室储存于–40 ℃的冰箱待测。进行测定时,取冷冻鲜样 0.2 g 于 20 mL 95%的乙醇溶液中研磨进行萃取,萃取液用于叶绿素含量的分析,Chla、Chlb 和 Car 分析参考,Chls 和 Chla/b 则通过计算获得。
1.3.4 叶面积指数、干物质积累和叶片 C、N 含量测定于大豆出苗后第 30、36 和 43 天,每处理小区选择 12 株取样,用蒸馏水清洗后,区分成根、茎和叶,用叶面积扫描仪扫描叶片,并采用图像分析软件 Scion Image 计算叶面积,叶面积指数根据种植密度和叶面积计算获得。根、茎和叶于 105 ℃杀青,80 ℃烘干至恒重,然后用电子天平(BP221S, Germany)称重,生物量和根冠比通过计算获得。称重后的部分叶片用小型高速粉碎机粉碎,过孔径 0.246 mm 筛,用于全碳、全氮含量的测定。全碳含量采用 K2Cr2O7氧化还原滴定法,全氮含量采用凯氏定氮法,碳氮比(C/N)为全碳含量与全氮含量的比值。
1.4 统计分析
数据的统计分析应用 SPSS 软件(version 15, SPSS, Chicago, IL, USA)进行。本研究中实验数据用单因素方差分析(one-way ANOVA)检验处理间差异的显著性,不同的字母表示差异显著(P < 0.05),相同字母表示差异不显著(P > 0.05)。不同参数之间的相关性采用 Pearson 相关矩阵进行分析。
2、 结果和分析
2.1 玉米-大豆套作对大豆幼苗生长微环境的影响
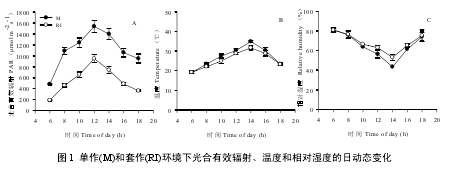
随全天时间变化,套作(RI)和单作(M)种植条件下光合有效辐射(PAR)和温度(TEMP)均呈先增加后降低的趋势,而湿度(RH)则表现为与 TEMP 变化相反的趋势(图 1)。6:00—18:00 时间内,RI 条件下 PAR 均显著低于 M (P < 0.01),平均下降程度达 52.5%;但 TEMP 和 RH 差异不显著(P > 0.05) (图 1)。这表明 RI 条件下的玉米遮荫将改变大豆幼苗的生长环境,显著降低了大豆幼苗生长发育所必须的可利用光资源。
2.2 玉米遮荫对大豆幼苗叶片叶绿素含量的影响
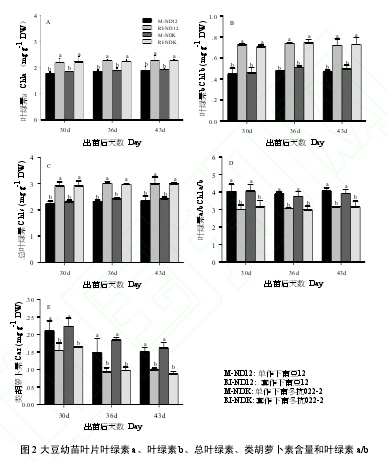
玉米遮荫能够显著提高大豆叶片 Chla、Chlb 和 Chls 的含量(图 2)。与 M 相比,RI 条件下,Chla、Chlb和 Chls 均显著升高(P < 0.05),2 个大豆品种 ND12 和 NDK 的 Chla 分别升高了 22.1%和 18.1%,Chlb 则分别升高了 58.4%和 49.4%,Chls 分别升高了 29.3%和 24.4%。这表明,玉米遮荫诱导了大豆幼苗叶片合成更多的 Chlb 来吸收更多的散射光,提高光的吸收率。而 Chla/b 和 Car 则表现为 RI 低于 M,且差异显著(P< 0.05)。2 个大豆品种在 3 次测定时均表现一致,但品种间无显著差异(图 2)。
2.3 玉米遮荫对大豆幼苗叶片 C、N 的影响
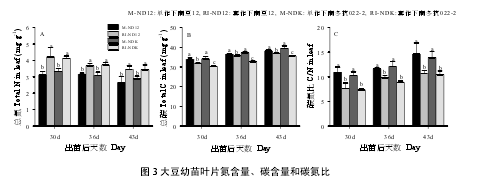
玉米遮荫显著提高了大豆幼苗叶片全氮(TON)含量(P < 0.05)(图 3A),这表明,玉米遮荫条件下,大豆幼苗将通过增加叶片氮含量来补偿因光照不足引起的光合速率下降。RI 条件下,叶片全碳(TOC)含量和碳氮比(C/N)均显著低于 M(P < 0.05)(图 3B,C),南冬抗 022-2(NDK)品种叶片 TOC 的下降程度更大(11.1%),显著高于南豆 12(ND12)(3.8%)(P < 0.05)。
2.4 玉米遮荫对大豆幼苗气体交换和叶绿素荧光参数的影响
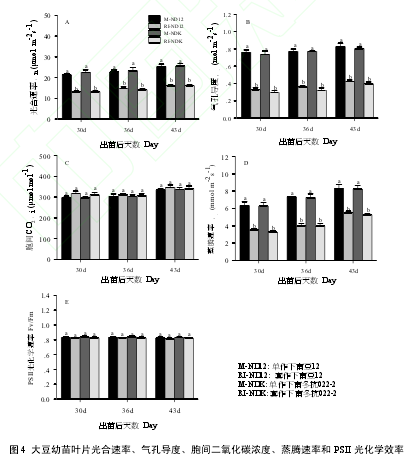
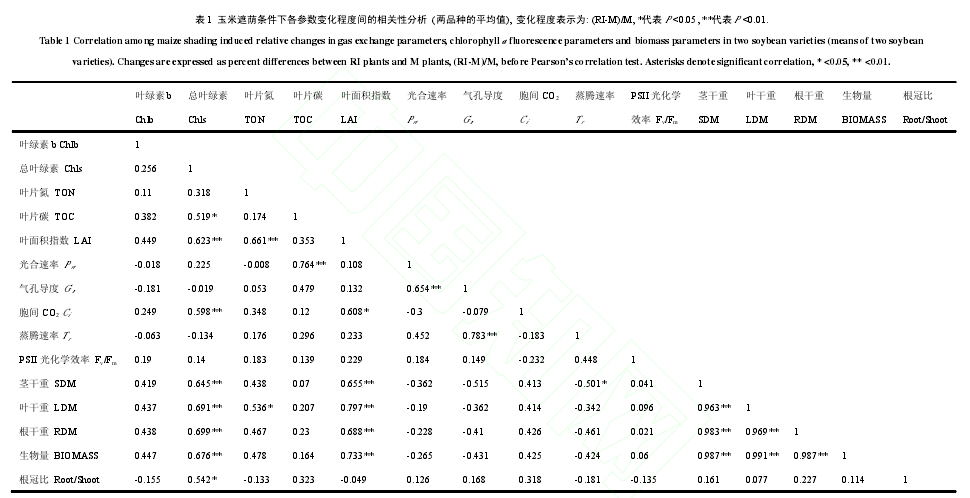
套作环境下,大豆幼苗叶片净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)受玉米遮荫的影响,均显著低于单作(P < 0.05)(图 4A,B,D),平均下降程度分别为 37.9%、54.2%和 42.4%,品种间无显著差异(P > 0.05),且3次测定时表现一致,玉米遮荫所导致的Pn的下降程度与Gs和TOC的下降程度呈极显著的正相关关系,相关系数分别为 0.654 和 0.764(表 1)。而胞间二氧化碳浓度(Ci) (图 4 C)和 PSII 光化学效率(Fv/Fm) (图 4E)在净作和套作条件下,2 品种在 3 次测定时均未表现出显著差异(P > 0.05)。
2.5 玉米遮荫对大豆幼苗叶面积指数的影响
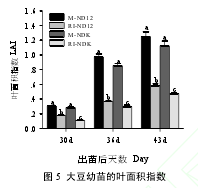
由图 5 可以看出,M 件下,2 大豆品种幼苗的叶面积指数(LAI)无显著差异,玉米遮荫显著降低大豆幼苗的 LAI,降低程度平均高达 57.1%。套作环境中 NDK 幼苗的 LAI,3 次测定值分别为 0.11、0.28 和 0.47,显著低于 ND12(0.17、0.37 和 0.58)和单作条件下的 LAI(0.29、0.91 和 1.19),表明 ND12 的耐荫性优于 NDK。
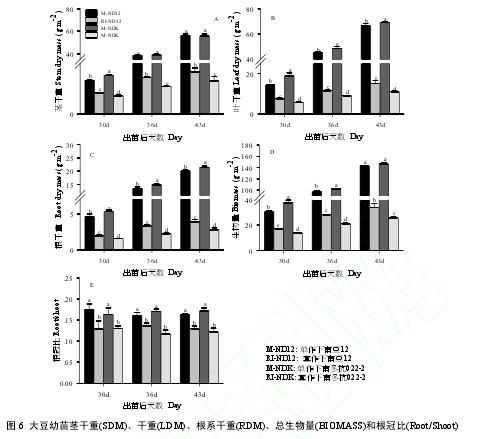
2.6 玉米遮荫对大豆幼苗干物质积累与分配的影响
由图 6 A 可以看出,RI 环境中,玉米遮荫显著降低了大豆幼苗的茎秆干重(SDM)(P < 0.05),且 NDK的降低程度(69%)显著高于 ND12(59%)。不同种植模式下,不同大豆品种的叶片干重(LDM)、根系干重(RDM)和总生物量(BIOMASS)存在显著的差异,具体表现为:M-NDK > M-ND12 > RI-ND12 > RI-NDK,玉米遮荫导致的 LDM、RDM 和 BIOMASS 的下降程度,NDK(78.5%、81.9%\和 75.3%)显著高于 ND12(66.2%、71.3%和 64.1%)(P < 0.05)(图 6 B,C,D),表明 ND12 的耐荫性优于 NDK。玉米遮荫所导致的 BIOMASS 下降程度与 LAI、SDM、LDM、RDM 的下降程度和 Chls 含量的变化程度存在极显著的正相关关系(表 1),而与 Pn不存在显著的相关关系。玉米遮荫还显著降低了大豆幼苗的根冠比(Root/Shoot)(P < 0.05),但品种间差异不显著(图 6 E)。
3、 讨论
间套作作为一种基于生物多样性的可持续农业发展模式,具有多种重要的生态服务功能。然而高位作物(玉米)的遮荫将会改变低位作物(大豆)的微生长环境,进而影响到系统的生产力,因此理解低位作物对系统微环境变化的适应机理,实现共生作物和谐发展和双高产是间套作模式推广和发展的关键。本研究发现,在玉米-大豆套作模式中,玉米遮荫降低了抵达大豆冠层的光合有效辐射(PAR),下降程度高达52.5%(图 1 A),这将引起大豆幼苗 Pn下调,同时 Gs和 Tr下降(图 4),与前人的研究结果一致。光合作用是物质代谢和能量转化的最初源泉,受植物自身生理特性和环境的共同影响。光环境对植物的直接作用就是光合作用,而关于荫蔽引起光合速率下降的原因国内外研究者存在一定的争议,有研究认为是气孔限制引起。也有研究认为是非气孔限制引起,主要是包括叶片叶绿素和可溶性蛋白含量降低导致捕获光合电子的能力下降,PSII 损伤导致电子传输能力降低或叶绿体的结构、数目和功能下降所引起。还有研究认为是气孔限制和非气孔限制共同作用的结果引起。本研究发现套作条件下,Pn的下降程度与 Gs的下降存在显著的相关关系(表 1),这表明玉米遮荫引发的大豆幼苗叶片气孔导度降低可能是引起光合速率下降的一个原因。另外,与单作相比,套作条件下大豆幼苗叶片 Ci和 Fv/Fm 无显著的差异(图4),这意味着玉米遮荫并未造成 PSII 受到损伤,Pn的下降可能由 CO2同化过程中的能量不足引起,而与光反应过程无关。因此,RI 环境中的玉米遮荫导致的 Pn降低可能是由气孔限制和 CO2同化过程中的能量不足共同引起,对于其具体的影响过程还有待于更深入的研究。
外界环境发生变化时,植物将会改变自身形态、结构和生理生化特性来适应环境,以便维持最佳的生长发育水平。弱光环境中,植物叶片将会调整自身结构和生理特征来吸收利用更多的光来维持生长发育,如叶片变薄、叶面积变大、光合色素含量增加、叶片 N 含量升高等。本研究中套作荫蔽环境会引起大豆幼苗叶片 Chla、Chlb、Chls(图 2)和 TON 含量(图 3)升高来吸收更多的散射光提高光能的捕获能力。然而,叶片 TOC 含量、LAI 和 LDM 的降低则使套作环境下大豆幼苗 Chlb 和 TON 含量升高所引起的光能捕获能力增强并不能补偿玉米遮荫所导致的光截获面积降低而引起的光截获量下降。玉米遮荫引起的叶片 TOC 含量的下降程度与 Pn的下降程度存在极显著的相关关系(表 1) ,说明 TOC 含量的下降可能主要是由玉米遮荫导致的大豆幼苗 Pn降低所引起。而 LAI 和 LDM 的下降可能主要是由弱光环境下大豆幼苗叶片形成的数量较低所引起。有研究表明,叶面积指数和叶片干物质重的变化是评价弱光环境中植物光敏感性的重要指标,本研究也发现,套作环境中光敏感型大豆品种 NDK 的 LAI 和 LDM 显著的低于耐荫大豆品种 ND12,因此,大豆叶片 LAI 和 LDM 可作为判断套作环境下大豆光敏感性差异的表观特征。
干物质积累是植物同化和固定 CO2能力的最直接表现,而弱光引起的植物干物质积累量变化的差异是直接反应物种间光敏感性的重要指标。套作荫蔽环境中,光敏感型大豆品种 NDK 的 SDM、RDM 和BIOMASS 下降程度显著高于耐荫性品种 ND12 (图 6),表明 NDK 的生长发育和 CO2固定能力受荫蔽胁迫程度更大,ND12 更适合应用于玉米-大豆套作模式。有研究表明,作物 95%以上的干物质来源于光合同化物,光合速率和光吸收面积是决定干物质形成的两个最重要方面。本研究发现,套作环境中大豆干物质积累下降可能主要是由 LAI 下降导致的 LDM、SDM 和 RDM 降低所引起(表 1), NDK 在套作环境下较低的叶面积指数可能是导致其生物量下降程度较大的主要原因。此外,玉米遮荫导致的大豆幼苗根冠比(Root/Shoot)下降(图 6 E)则可能意味着套作环境中碳同化物更多的被分配到地上部来最大程度地获取光照资源。
4、 结论
玉米-大豆套作模式中高位作物(玉米)遮荫会引起低位作物(大豆)幼苗光合作用显著下降,具体表现为:叶片 Pn、Gs、Tr、LAI、TOC、SDM、LDM、RDM 和 BIOMASS 显著下降;而与光吸收相关的叶片光合色素 Chla、Chlb、Chls 以及 TON 含量却显著上升。光合速率 Pn的下调主要是由气孔限制和 CO2同化过程中的能量不足共同引起,而干物质积累的降低则主要是由 LAI 降低所引起。LAI、SDM、LDM、RDM 和BIOMASS 可作为生产和育种方面评价大豆品种耐荫性差异的良好指标。耐荫性品种 ND12 更适合应用于西南地区玉米-大豆套作模式中,深入研究其遗传和生理生化特性对选育更适合于玉米-大豆套作模式的大豆品种具有重要的意义。
致谢:感谢匿名审稿专家提出的宝贵修改意见;感谢许宏博士对文章撰写的帮助。





